Bot. Bull. Acad. Sin. (2002) 43: 291-298
Sasikiran et al. Proteinaceous enzyme inhibitor levels in sweet potato
Proteinase and alpha-amylase inhibitors of sweet potato: Changes during growth phase, sprouting, and wound induced alterations
K. Sasikiran, M.R. Rekha, and G. Padmaja*
Central Tuber Crops Research Institute, Thiruvananthapuram, 695017, Kerala, India
(Received May 8, 2002; Accepted July 12, 2002)
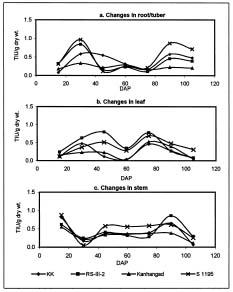
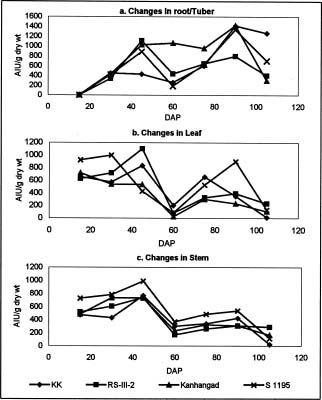
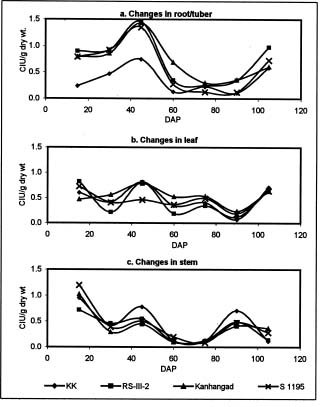
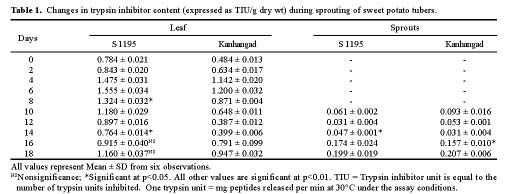
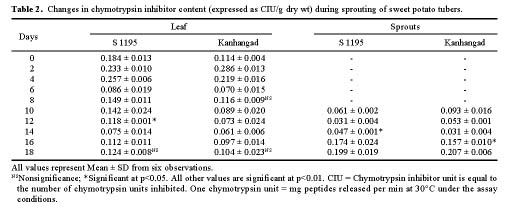
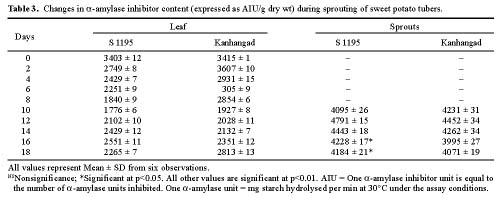
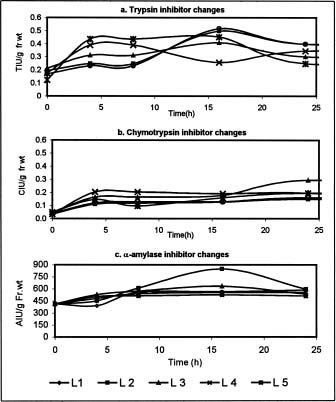
Abstract. Changes in proteinase (trypsin and chymotrypsin) and a-amylase inhibitors were monitored during the growth phase and sprouting of sweet potato, with a view to establishing the biological role of these secondary plant principles. Although the trypsin inhibitor (TI) activity of tuberising roots of sweet potato was very low in four genotypes at 15 days after planting (DAP), a steep increase was observed during the tuber-bulking phase, indicating a possible role for the inhibitor in regulating plant proteinases. In contrast, the chymotrypsin inhibitor (CI) activity decreased from 40-90 DAP, indicating an inconsequential role in protease regulation. With initiation of tuber bulking, the a-amylase inhibitor (AI) increased sharply in three sweet potato genotypes, and after completion of the bulking phase, the activity came down sharply at 105 DAP. A pronounced role for regulation of endogenous a-amylases by the AI of sweet potato could be postulated from the study. The TI activity increased in the tubers until the sixth day of planting in vermiculite, and it decreased with the emergence of sprouts, indicating the possibility of proteolytic cleavage at the time of sprouting. Amylase inhibitor activity continuously decreased until sprouting, indicating the role of these proteins as N-reserve proteins in sweet potato. A significant increase in proteinase inhibitor levels in all the sweet potato leaves was observed within four hours of artificial wounding. The rise in inhibitor levels was more evident in leaves far off from the wounded leaf. The fact that proteinase and amylase inhibitors of sweet potato are wound-inducible points to a major defensive role for them. This information on the proteinase inhibitor levels in sweet potato leaves and vines and their change with plant growth is of interest, especially due to their use as animal feed.
Keywords: a-Amylase inhibitor; Chymotrypsin inhibitor; Growth phase; Ipomoea batatas; Sprouting; Trypsin inhibitor; Wounding.
Abbreviations: AI, a-amylase inhibitor; TI, trypsin inhibitor; CI, chymotrypsin inhibitor; DAP, days after planting.
Introduction
Recent developments in the field of proteinase and amylase inhibitors and the increasing use of these phytochemicals in industry and pharmacy have led researchers to inquire closely into their exact biological functions. Their role as natural plant defense proteins has attracted a lot of attention (Farmer and Ryan, 1990; Gatehouse et al., 1986). These inhibitors are believed to make plants less palatable, even lethal, to insects, thus conferring some selective advantage to the plants. Several workers have reported a decrease in proteinase inhibitor activity with germination (Ambekar et al., 1996; Hobday et al., 1973). The transient nature of potato chymotrypsin inhibitor I was reported by Ryan et al. (1968). The changes in trypsin inhibitor activity in the vegetative tissues of sweet potato plants were studied by Zhang and Corke (2001). Mulimani et al. (1994) reported a decrease in a-amylase inhibitor (AI) activity with germination in 28 varieties of chickpea. Reports on the alpha amylase inhibitor
changes in plants are scanty, except those indicating alpha amylase accumulation in the seeds of cereal crops during seed formation (Mundy, 1984; Pace et al., 1978). The wounding of the lower terminal leaflets of tomato plants by Colorado potato beetles has been reported to lead to the accumulation of proteinase inhibitor in both wounded and non-wounded leaves (Green and Ryan, 1972). Artificial wounding by heat or even crushing with a file can mimic the action of an insect bite and stimulate the production of the inhibitor (Green and Ryan, 1972; Hilder et al., 1987). Yeh et al. (1997) found that transgenic tobacco plants carrying sweet potato TI genes were resistant to the tobacco cut worm, Spodoptera litura.
We first reported the presence of proteinase and a-amylase inhibitors in a large number of sweet potato genotypes (Sasikiran et al., 1999; Rekha et al., 1999). Our later studies highlighted the inhibitor potential of purified protease and a-amylase inhibitors from five sweet potato genotypes on the digestive enzymes of root crop storage pests (Sasikiran, 2000; Rekha, 2000). In this paper, we report the changes in the trypsin, chymotrypsin, and a-amylase inhibitors in sweet potato during growth phase, sprouting, and wounding.
*Corresponding author. Tel: 0091-471-598851; Fax: 0091-471-590063; E-mail: g_padmaja1@rediffmail.com