Bot. Bull. Acad. Sin. (2003) 44: 1-11
Chao et al. A new rice transposable element
TEOS1, a novel transposable element family from Oryza sativa
Ya-Ting Chao, Chun-Lin Su, Teh-Yuan Chow, Pei-Fang Lee, Chun-I Chung, Jiann-Jang Huang, Su-Mei Liu, and Yue-Ie C. Hsing*
Institute of Botany, Academia Sinica, Taipei, Taiwan 11529
(Received June 18, 2002; Accepted August 16, 2002)
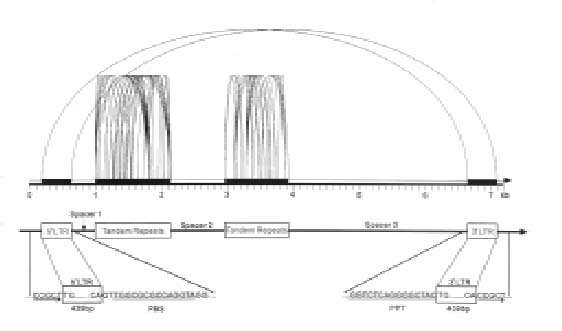
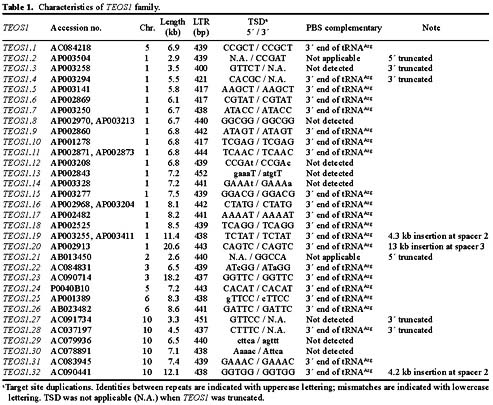
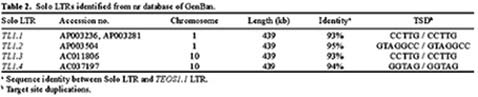
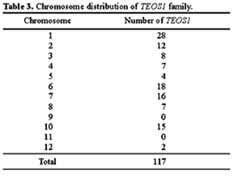
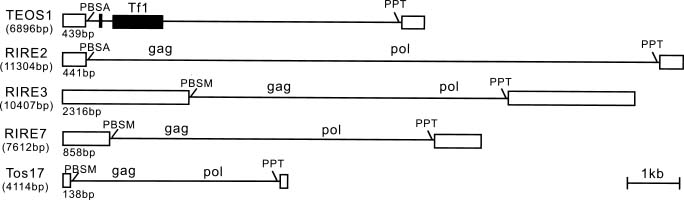
Abstract. We have uncovered a novel group of transposable elements TEOS1, (Transposable Element of Oryza sativa) by repeat mining of rice genome sequence data. We first identified a TEOS1 element, 6,896 bp in length, from a PAC clone of rice chromosome 5. The presence of long terminal repeats (LTRs) and other diagnostic features, such as a primer-binding site (PBS) and a polypurine tract (PPT), indicated that TEOS1 is a group of LTR retrotransposon. The internal domain of TEOS1 containing two arrays of tandem repeats and a predicated Tf1gene was characteristically distinct from those of other LTR retrotransposons previously described. TEOS1 elements identified were flanked by 5-bp direct repeats; some identified as solo LTRs were flanked by 5-7 bp direct repeats. The PBS sequence was homologous to the 3´ end region of tRNAArg. The deduced amino acid sequence of Tf1 is dissimilar to that of any known mobility related proteins. TEOS1 is probable to be trans-activated by other retrotransposons in the rice genome. We have estimated that there are about 300 TEOS1 elements per haploid rice genome. The presence of repeats in TEOS1 offers the potential to form secondary structure during replication and transcription, and to serve as a hot spot for transposon insertion, augmenting the dynamics of molecular interactions of the genomic sequences.
Keywords: Oryza sativa; Tandem repeat; TEOS1; Transposable element.
Abbreviations: GSS, Genomic survey sequence; HTG, Unfinished high throughput genomic sequence; LINE, Long interspersed repetitive element; LTR, Long terminal repeat; MITE, Miniature inverted repeat transposable element; PBS, Primer binding site; PPT, Polypurine tract; SINE, Short interspersed repetitive element; TE, Transposable element; TEOS, Transposable element of Oryza sativa; U5, Unique 5´ RNA.
Introduction
Transposable elements (TE) are divided into two main classes according to their mechanism of transposition. Retrotransposons, termed as Class I elements, transpose through an RNA intermediate that is converted into extrachromosome DNA and integrated into the genome. The replication mode could increase their copy numbers rapidly and thereby greatly increase the size of the host genome (Kumar, 1996; SanMiguel and Bennetzen, 1998). In some plants, retrotransposons comprise over 50% of the nuclear DNA content (Pearce et al., 1996; SanMiguel et al., 1996; Suoniemi et al., 1996; Kossack and Kinlaw, 1999). According to the structural organization and the encoded ORFs, retrotransposons could be further typed as copia, gypsy, and the non-LTR retrotransposon. The copia and gypsy type retrotransposons are characterized by the flanking long terminal direct repeats (LTRs), and distinguished by the order of the domains in their polyprotein region.
Class I elements have been characterized in a wide range of plant taxa. The first Class I element identified in plants is Tnt1 in tobacco (Grandbastien et al., 1989). Oth
ers plant elements include Tto1-Tto3 in tobacco (Hirochika, 1993), BARE-1 in barley (Suoniemi et al., 1996), Bs1 (Jin and Bennetzen, 1989) and Opie-1 in maize (SanMiguel et al., 1996), soybean SIRE-1 in soybean (Laten et al., 1998), and Athila (Pelissier et al., 1995) and Tal 1 (Wright et al., 1996) in Arabidopsis.
Due to its small genome size of 430Mb, the smallest among the cereal grasses (Arumuganathan and Earle, 1991), and concerted efforts in its sequence analysis, several retrotransposon families have been well identified in the rice genome. These include copia type Ty1 (e.g. Tos 1-20) (Hirochika et al., 1992, 1996), gypsy type Ty 3 (e.g. RIRE3, RIRE7) (Kumekawa et al., 1999, 2001), and SINE (e.g. p-SINE1) (Mochizuki et al., 1992; Motohashi et al., 1996). Cell culture-induced activation of Tos17 in rice has been applied to insertional mutagenesis analysis (reviewed in Hirochika, 2001). For example, the zeaxantin epoxidase gene has been tagged using Tos17 insertion (Agrawal et al., 2001). The feasibility of using Tos17 in the PCR-screening strategy was demonstrated by screening for a mutant of the homebox gene OSH15 (Sato et al., 1999). Previous studies using computer-based sequence similarity searches have revealed the predominance of MITE elements in rice (Bureau and Wessler, 1994; Bureau et al., 1996; Mao et al., 2000; Turcotte et al., 2001). Many TEs are also identified in rice BAC end sequences (Mao et al., 2000) or genomic sequence (Turcotte et al., 2001).
*Corresponding author. Tel: 886-2-27899590 ext. 312; Fax: 886-2-27827954; E-mail: bohsing@gate.sinica.edu.tw