Bot. Bull. Acad. Sin. (2003) 44: 19-24
Lo et al. Actin-like protein filaments inside mitochondria
The presence of actin-like protein filaments in higher plant mitochondria
Yih-Shan Lo, Yuh-Tai Wang, Wann-Neng Jane, Lin-June Hsiao, Long-Fang O. Chen, and Hwa Dai*
Institute of Botany, Academia Sinica, Taipei, Taiwan 11529, Republic of China
(Received May 2, 2002; Accepted July 25, 2002)
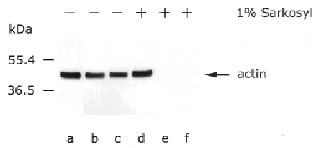
Abstract. In this report we demonstrate that a 42 kDa actin-like protein filament may exist in mung bean seedling mitochondria. The results of a fine structure study, immunogold localization, immunoblot, fluorescent microscopy, and flow cytometric analyses all point to the existence of this actin-like protein inside mung bean mitochondria.
Keywords: Actin-like protein; Plant mitochondria.
Introduction
The cytoskeleton in eucaryotic cells has been well studied. It determines the cell shape and directs the movement of various entities in the cytoplasm. The transport action of mitochondria from one place to another in cytoplasm, controlled by the actin cytoskeleton, has also been well characterized (Kuznetsov et al., 1992; Simon et al., 1995). However, very little is known about how mitochondria keep their shape or how newly synthesized mitochondrial gene-encoded proteins are transported to their final destination for function. Most studies on mitochondrial biogenesis in the past two decades have focused on the import pathway of nucleus gene-encoded proteins into mitochondria (Baker and Schatz, 1991; Pfanner et al., 1992; Fujiki and Verner, 1993).
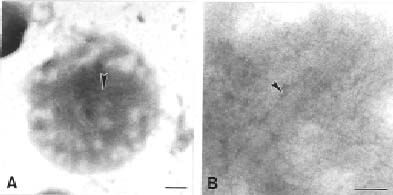
The existence of actin-related protein inside the chloroplast and mitochondria has been suggested before (McCurdy and Williamson, 1987; Murgia et al., 1995). That the F-actin is probably present in organelle dividing rings was suggested by Kuroiwa et al. (1995). Miyagishima et al. (2001) proved that a 5 nm novel filament was involved in the plastid-dividing ring. The tubulin homologous protein-FtsZ, an essential cell division protein in bacteria, was well studied (Bi and Lutkenhaus, 1991; Erickson, 1995, 1997). Osteryoung and Vierling (1995) demonstrated that cpFtsZ, a homologue of tubulin protein, can be imported into the chloroplast of Arabidopsis. Mitochondrial FtsZ in a Chromophyte Alga was also reported recently (Beech et al., 2000). Most importantly, an actin-like protein (MreB) was found in prokaryotes (Jones et al., 2001; Van den Ent et al., 2001). In this report, we found about 5 nm of filamentous structures inside mitochondria. Immunoblot, immunolocalization, fluorescent analyses, and flow
cytometry all showed that 42 kDa actin-like protein filaments are present in mung bean seedling mitochondria.
Materials and Methods
Mitochondria, Mitoplast Isolation and EM Examination
Mitochondria were isolated from four-day-old etiolated mung bean seedlings (TN-5) with M-1 buffer as described previously (Leaver et al., 1983; Dai et al., 1991; Cheng and Dai, 2000). They were then treated with digitonin following the methods of Kfzlfwski and Zagorski (1998). Mitochondria were dissolved in medium B (0.6 M sorbitol, 0.2% BSA, 10 mM Tris-maleate, pH 7.0) at a protein concentration of 40 mg/ml. The digitonin concentration for mitochondria treatment was 0.12 mg/ml followed by a pre-optimized analysis. Mitoplasts were further purified with a 40-65% sucrose gradient buffered with 10 mM Tris-maleate (pH 7.0) by centrifuging at 100,000 g at 4°C for 90 min. Whole-mount EM analysis on mitoplast was carried out by fixing the sample with 1% paraformaldehyde followed by 1% uranyl acetate (pH 4.5) staining for 1-2 min.
Immunolocalization Analysis of Mitochondria in Situ
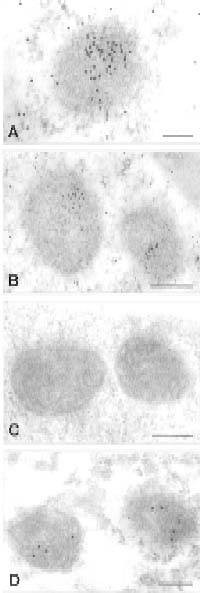
The four-day-old mung bean tissue was fixed in 10 mM pipes (pH 7.2), containing 2.5% paraformaldehyde and 1.25% glutaraldehyde at 4°C overnight. Dehydration was done after five washings in the same buffer. Infiltration and polymerization were carried out in Lowicryl K4M resin (Roth et al., 1981). Thin tissue sections were labeled by sequential incubations with monoclonal anti-actin antibody (Boehringer Mannheim, C4 clone) followed by reaction with a secondary antibody, goat-anti-mouse antibody conjugated with 20 nm gold. The maize mitochondria specific monoclonal antibody, PM028 (against a yet unidentified maize mitochondrial protein with no cross
*Corresponding author. Fax: 886-2-2782-7954; E-mail: bodaihwa@gate.sinica.edu.tw