Bot. Bull. Acad. Sin. (2003) 44: 25-30
Huang et al. Developmental phase reversal of Sequoia sempervirens
Association of mitochondrial plasmids with rejuvenation of the coastal redwood, Sequoia sempervirens (D. Don) Endl.
Li-Chun Huang1, Teh-Yuan Chow1, Tsung-Che Tseng1, Ching-I Kuo1, Su-Mei Liu1, Moon-Geok Ngoh1, Toshio Murashige1, and Hao-Jen Huang2,*
1Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan
2Department of Biology, National Cheng-Kung University, Tainan, Taiwan
(Received May 9, 2002; Accepted August 16, 2002)
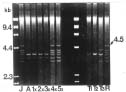
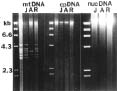
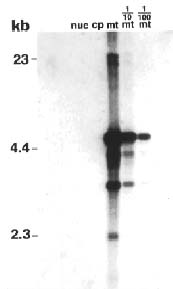
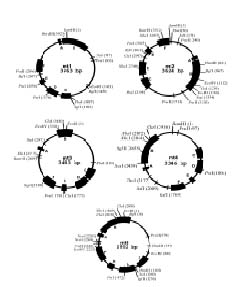
Abstract. Repeated grafting of shoot apices from mature Sequoia sempervirens (D. Don) Endl. trees onto young seedling shoots in vitro causes emergence of rejuvenated, or phase-reversed, scion shoots. The phase reversal is indicated by a restored rooting competence and renewed vigor of roots and shoots. We earlier reported of restriction fragment length polymorphism between juvenile or rejuvenated and adult shoot mtDNA (mitochondrial DNA). This investigation of undigested mtDNA disclosed that S. sempervirens shoots contained at least six small mtDNA molecules, four of which were uniquely associated with juvenile and rejuvenated shoots. The small molecules remained observable in continuously subcultured shoots. Results of cloning and sequencing indicated they are circular in form; we thus, called them plasmids. Significant sequence homology, 38 to 56%, was found among the molecules, indicating highly conserved regions and possibly common origin. No similar phase-associated relationships were observed for unrestricted cpDNA (chloroplast DNA) or nucDNA (nuclear DNA). The plasmids hybridized with the larger, electrophoretically less mobile mtDNA, but not with cpDNA or nucDNA, suggesting their origin in the master circle mtDNA. The distinctness of these plasmids has remained unchanged after more than 20 years of subculturing.
Keywords: Mitochondrial plasmids; Rejuvenation; Sequoia sempervirens.
Introduction
Development of the sexually reproductive mature adult plant normally begins with a strictly vegetative juvenile phase. The maturation, or phase change, is completed within weeks among annuals, but can span several years among perennials. In trees, the process is frequently accompanied by ancillary morphological and physiological traits, most commonly a loss of competence for adventitious rooting and of overall vigor. Since a plant's developmental phase is determined in its shoot apical meristems, reversing the phase of the meristems should result in emergence of rejuvenated shoots. Indeed, new growths with reversed phases have been obtainable variously, including through gibberellin (Rogler and Hackett, 1975) or cytokinin (Bouriquet et al., 1985; Franclet et al., 1987) applications to plants; through continuous subculturing of shoots, especially in cytokinin-containing media (Fouret et al., 1885; Lyrene, 1981; Walker, 1986; Brand and Lineberger, 1992); and through repeatedly grafting shoot apices from mature trees onto juvenile rootstocks in vivo (Doorenbos, 1954; Muzik and Cruzado, 1958) or in vitro (Monteuuis, 1986; Pliego Alfaro and Murashige, 1987; Huang et al., 1992a, 1992b; Ewald and Kretzschmar, 1996; Revilla et al., 1996).
Our earlier investigation of S. sempervirens disclosed mtDNA restriction fragment length polymorphism between juvenile and adult shoots (Huang et al., 1995). Juvenile and rejuvenated shoots contained 4.0 and 3.6 kb BamHI-restricted fragments that were absent in adult shoots. We now report on finding distinctive mitochondrial plasmids in juvenile and rejuvenated shoots.
Materials and Methods
Tissue Culture
Continuously cultured Sequioa sempervirens (D. Don) Endl. shoots from freshly germinated seedlings served as the juvenile rootstocks, and those from mature trees provided the adult shoot meristems. The continuous cultures were initiated from terminals of juvenile and adult shoots and from scion-growth remnants following regrafting of shoot tips. One- centimeter long terminals were subcultured at 6-wk intervals in nutrient media containing Murashige and Skoog salts (Murashige and Skoog, 1962), 3% sucrose, 0.2% gelrite, and, in µM: 555 i-inositol, 3 thiamine·HCl, 2.4 pyridoxine·HCl, 4.1 nicotinic acid, and 26.6 glycine. Grafting was done by inserting the obliquely cut base of a 1.5-cm long shoot terminal into a longitudinal incision made in a rooted 1-cm tall juvenile stem segment. Regrafting was done at 8-wk intervals. Shoot cultures and grafted plants were maintained at 27°C and under 16-h daily exposure to 22.5 µmol m-2 s-1 cool-white
*Corresponding author. Tel: +886-6-2757575 ext. 65539; Fax: +886-6-2742583; E-mail: haojen@mail.ncku.edu.tw