Bot. Bull. Acad. Sin. (2003) 44: 31-35
Huang et al. Phase change and photosynthesis in Sequoia sempervirens
Photosynthetic potentials of in vitro-grown juvenile, adult, and rejuvenated Sequoia sempervirens (D. Don) Endl. shoots
Li-Chun Huang, Jui-Hsi Weng, Chiu-Hui Wang, Ching-I Kuo, and Yuh-Jang Shieh*
Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 11529
(Received June 7, 2002; Accepted November 7, 2002)
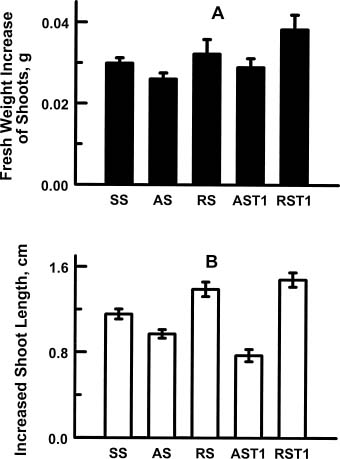
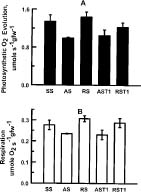
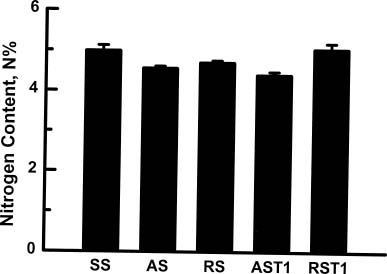
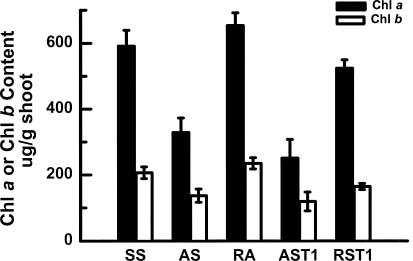
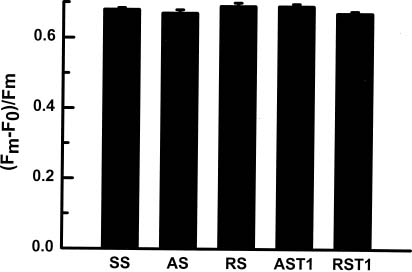
Abstract. In vitro shoot tips of Sequoia sempervirens (D. Don.) Endl.including a juvenile, two adult, and two rejuvenated adult cloneswere examined for differences in basic physiological characteristics. Moisture contents were the same, around 85%, for all tissues regardless of origin. Growth rates, determined by fresh weight increase and shoot elongation, were higher for the juvenile and rejuvenated shoots. They correlated with higher total nitrogen contents. Juvenile and rejuvenated shoots also showed higher rates of photosynthesis and respiration, evidenced by faster O2 evolution and consumption. The photosynthetic rates were associated with more chlorophyll, especially chlorophyll a, in the juvenile and the rejuvenated shoots. Nevertheless, identical quantum efficiencies of photosystem II indicated the same photosystems were operating and with equal effectiveness in juvenile, adult, and rejuvenated tissues.
Keywords: Phase change; Photosynthesis; Respiration; Sequoia sempervirens; Total nitrogen content.
Introduction
Plant development normally begins with a strictly vegetative juvenile phase and culminates in the sexually reproductive adult phase. Although herbaceous annuals achieve phase change, or maturation, within weeks, in woody perennials (Franclet, 1983), especially trees, this phase change or maturation process can take several years and is associated with changes of different characteristics, e.g., loss of adventitious rooting competence, diminished vigor and growth rate, changes in phyllotaxy, shape and size of leaves, re-orientation of leaves from plagiotropism to orthotropism, and decreased thorniness depending on species. During the past several years we have been employing Sequoia sempervirens (D. Don.) Endl., the coastal redwood, as a model to characterize the juvenile and mature phases of plants through their biochemical and physiological differences, as well as to progress in the understanding of the underlying mechanism controlling phase change in trees. This species was chosen because its excised shoots can be easily cultured in vitro on a chemically defined medium without phytohormonal supplements. Also, a phase reversal resulting in emergence of juvenile shoots from adult shoots is readily achieved (Huang et al., 1992). Furthermore, the shoots of the two phases display distinct morphogenetic characteristics. Repeated grafting in vitro of adult shoot tips onto rooted shoot segments of juvenile seedlings eventually results in scion growths with juvenile characteristics, such as a higher capacity for adventitious rooting, vigorous growth, and plagiotropic stems (Huang et al., 1992). Differences in esterase and peroxidase
isozymes were observed between juvenile or rejuvenated and adult shoots (Huang et al., 1996). The rate of protein phosphorylation was also higher in the juvenile shoots (Kuo et al., 1995), compared to the adult ones. Although juvenile and rejuvenated S. sempervirens produced more ethylene per cultured shoot, the rate of ethylene emission per gram of tissue was found to be the same (Huang et al., 2000). Restriction fragment length polymorphism of mtDNA (mitochondrial DNA) was also pointed out between juvenile and adult phase shoots (Huang et al., 1995). In this report, phase change aspects in tissue-cultured Sequoia sempervirens are investigated with respect to the potential for photosynthesis, and this is complemented by information on respiration rates and nitrogen contents.
Materials and Methods
Tissues Analyzed
Five different origins of shootsjuvenile (SS), adult from two different trees (AS and AST1), and rejuvenated shoots (RS and RST1) from the two adult S. sempervirens were investigated. Stocks of juvenile shoots were initiated from seedlings germinated in vitro. The adult stocks were established by culturing shoot tips excised from trees that were at least 60 years old. Stock cultures of AS were initiated from shoots excised in 1976, and those of AST1 were established from another mature tree in 1994 (Huang et al., 2000). Stocks of rejuvenated shoots, RS and RST1, were derived from AS and AST1, respectively. Rejuvenated shoots were obtained by 5-times grafting of the shoot tips from the two mature trees onto rooted juvenile seedling segments in vitro. Shoots of all five were available in stock cultures, maintained by monthly subculturing on a medium containing MS (Murashige and Skoog, 1962) salts, 3%
*Corresponding author. E-mail: yjshieh@gate.sinica.edu.tw