Bot. Bull. Acad. Sin. (2003) 44: 113-122
Park et al. A test of hybrid origin of Paraixeris koidzumiana
Allozyme variation in Paraixeris: a test for the diploid hybrid origin of Paraixeris koidzumiana (Compositae)
Ki-Ryong Park1,*, Jae-Hong Pak2, and Bong-Bo Seo2
1Department of Biology, Kyung-Nam University, 449 Wolyoung-dong, Masan 631-701, Korea
2Department of Biology, College of Natural Sciences, Kyungpook National University, Daegu 702-701, Korea
(Received August 7, 2002; Accepted November 18, 2002)
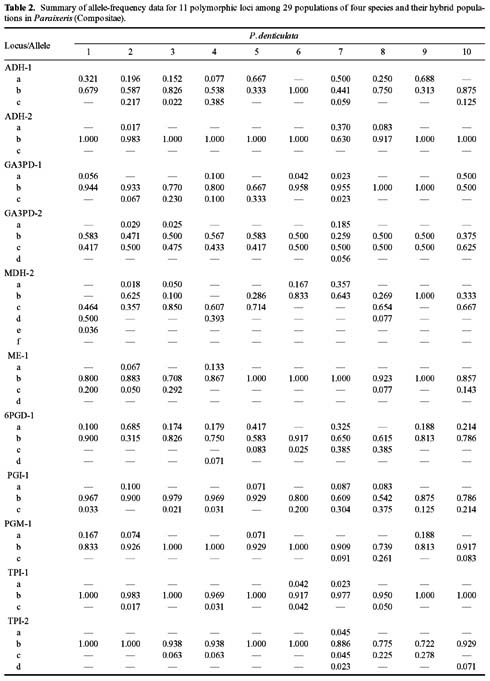
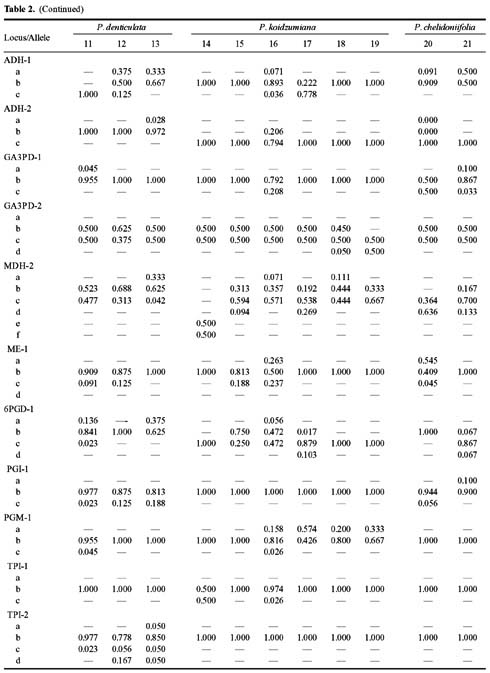
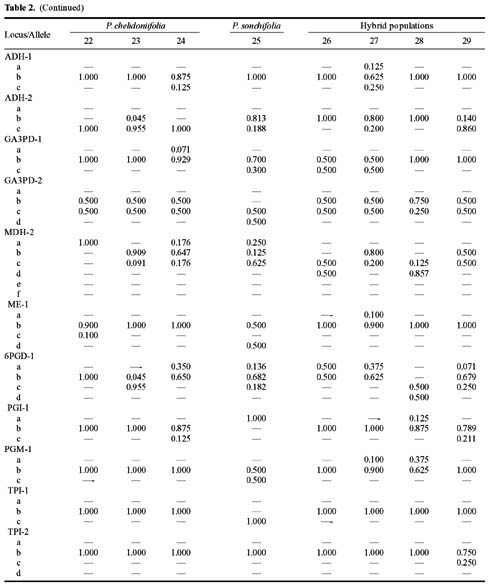
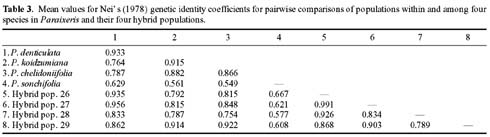
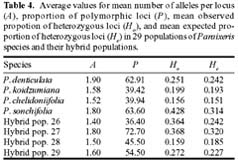
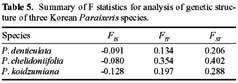
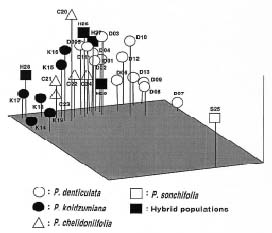
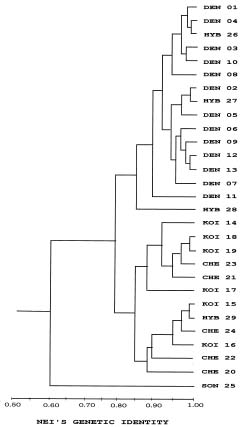
Abstract. Variation in isozyme patterns of four Korean Paraixeris species was used to test the hypothesis that P. koidzumiana is a hybrid derivative of P. chelidoniifolia and P. denticulata, or alternatively, of the progenitor P. chelidoniifolia-P. sonchifolia pair. Eleven loci from eight enzymes were examined for 29 populations of P. koidzumiana, P. denticulata, P. chelidoniifolia, and P. sonchifolia. Although recent RAPD and morphological data supported the hybrid origin of P. koidzumiana, isozyme evidence did not. No evidence of additivity was found in P. koidzumiana at the loci that significantly differentiate P. sonchifolia from other species. In addition, P. koidzumiana populations did not combine the marker alleles differentiating P. denticulata from P. chelidoniifolia. Recent divergence between P. koidzumiana and P. chelidoniifolia is strongly supported by the high genetic identity value and sharing of the same high frequency alleles at most loci. Our isozyme data largely support a conclusion that the unidentified individuals found in species sympatry are hybrid derivatives.
Keywords: Hybridization; Isozyme; Paraixeris koidzumiana.
Introduction
Natural hybridization is common in flowering plants and provides an important source of increasing genetic variation, creating novel lineages via stabilization of the hybrid derivatives and breakdown or reinforcement of isolating barriers (Levin, 1979; Rieseberg and Ellstrand, 1993; Wolfe and Elisens, 1993; Arnold, 1992, 1997; Morrell and Rieseberg, 1998; Nielsen, 2000). However, due to sterility or limited fertility in hybrid individuals, and the lack of molecular markers to detect parental taxa, diploid hybridization has been assigned only minor evolutionary importance (Arnold, 1992; Wolfe et al., 1998). Several recent studies of plant speciation suggest that new diploid species might arise rapidly through hybridization between genetically divergent species (Ungerer et al., 1998). In this case, genetic additivity of allozyme data has been useful in determining parental species (Gallez and Gottlieb, 1982). However, the detection of ancient hybrid origin may be difficult due to the parental taxa and its derivatives having had a chance to accumulate new genetic variation (Morrell and Rieseberg, 1998). Although many cases of recombinational speciation in the Far East Asian Paraixeris group have been proposed based on cytological and morphological studies (Ono, 1946, 1950, 1951), few have been tested with molecular markers.
The Korean endemic Paraixeris koidzumiana Kitam. (Kitamura, 1942; Pak, 1991) is a diploid (2n = 10) herbaceous biennial, restricted to Mt. Chiri, while the closely
related P. chelidoniifolia, P. denticulata, and P. sonchifolia are also diploid, but widely distributed in Korea, Japan, and Manchuria (Pak and Kawano, 1990). Since Kitamura (1942) described P. koidzumiana from the specimen of Koidzumi collected from Mt. Chiri, Lee (1979) proposed that P. koidzumiana originated from hybridization between P. sonchifolia and P. chelidoniifolia. This hybrid origin was originally proposed because of the intermediate leaf morphology between the two species. Paraixeris koidzumiana resembles P. chelidoniifolia in having pinnately divided leaves, but is similar to P. sonchifolia in having margined petioles that clasp at the base. However, a cytological study by Pak (1991) did not give support to P. sonchifolia being one of the parental species of P. koidzumiana. Additionally, the difference between the flowering periods of P. sonchifolia and P. chelidoniifolia argues against their hybridization (Tae et al., 2001). A recent molecular study using RAPD data supports the hybrid origin of P. koidzumiana from P. denticulata (Houtt.) Nakai and P. chelidoniifolia (Tae et al., 2001). In addition, at Mt. Chiri, where the above three species grow sympatrically, plants were discovered with leaves and inner-involucre numbers that appeared to be intermediate between P. chelidoniifolia and P. denticulata. Canonical variate analysis on the morphological characters suggested that the individuals were hybrids derived from P. chelidoniifolia and P. denticulata (Pak, unpublished data). The objective of the study was to determine, using isozyme analysis, whether P. koidzumiana and putative hybrid populations were derived through hybridization among P. chelidoniifolia, P. denticulata, and P. sonchifolia.
*Corresponding author. Tel: 82-55-249-2240; E-mail: park@kyungnam.ac.kr