Bot. Bull. Acad. Sin. (2003) 44: 141-150
Tsai and Wang Rice protein kinases phosphorylating sucrose synthase
Identification of rice manganese-dependent protein kinases that phosphorylate sucrose synthase at multiple serine residues
Zheng-Chia Tsai and Ai-Yu Wang*
Department of Agricultural Chemistry, National Taiwan University, No. 1, Section 4, Roosevelt Road, Taipei, Taiwan 106
(Received October 2, 2002; Accepted January 16, 2003)
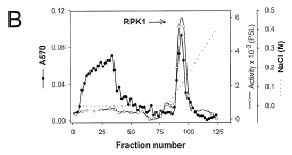
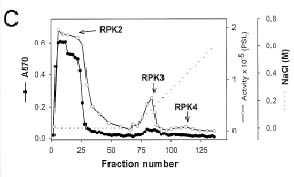
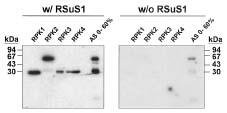
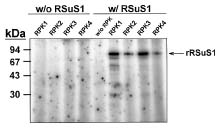
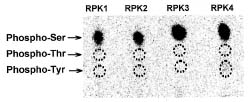
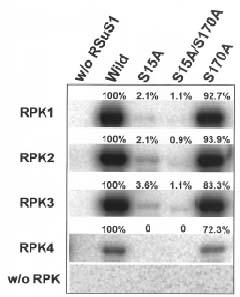
Abstract. Sucrose synthase from the etiolated seedlings of rice (Oryza sativa L. cv. Tainung 67) was phosphorylated both in vivo and in vitro. Four protein kinases that phosphorylated recombinant rice sucrose synthase 1 (RSuS1) in a Mn2+-dependent manner were partially purified and characterized from etiolated rice seedlings. These four kinases, designated as RPK1, RPK2, RPK3 and RPK4, are monomeric enzymes with apparent molecular masses of 34 kDa, 57 kDa, 30 kDa, and 30 kDa, respectively. Phosphoamino acid analysis of the 32P-labeled phosphorylated recombinant RSuS1 indicated that it was phosphorylated at serine residues by these four RPKs. RP-HPLC/ESI-MS analysis of the tryptic peptides of phosphorylated RSuS suggested that the serine residues in the tryptic peptides 13-LHSVR-17 and 168-HLSSK-172 were the target residues for phosphorylation. For confirmation of this finding, mutant recombinant RSuS1, S15A, S170A and S15A/S170A, were purified and subjected to phosphorylation by the four partially purified kinases. The results showed that both Ser15 and Ser170 residues were target residues for RPK1, PRK2 and PRK3 and Ser15 was the major phosphorylation site in RSuS1. Phosphorylation of RSuS1 may not occur exclusively at these two sites since weak phosphorylation of the double mutant protein S15A/S170A was also observed. Phosphorylation of the mutant S15A and S15A/S170A by RPK4 was undetectable, indicating that Ser15 was the only target residue for this kinase.
Keywords: Manganese-dependent protein kinases; Rice; Sucrose synthase.
Introduction
Sucrose synthase (UDPG: D-fructose 2-glucosyl transferase, SuS) was first described by Cardini et al. (1955) and has been characterized in various plant species. The enzyme catalyzes the conversion of sucrose and UDP into fructose and UDPG. Although the reaction is readily reversible, it is thought that SuS functions primarily in the direction of sucrose degradation to provide sugar nucleotides for complex saccharides synthesis (Chourey and Nelson, 1976; Amor et al., 1995; Déjardin et al., 1997; Chourey et al., 1998).
In most plants, SuS is encoded by two or three genes that are spatially and temporally regulated and are differentially modulated by the sugar level, anaerobiosis (Zeng et al., 1998; Wang et al., 1999; Winter and Huber, 2000 and references therein; Barratt et al., 2001; Carlson et al., 2002), and osmotic stress (Déjardin et al., 1999). In potato, SnRK1 (SNF1-related protein kinase) activity is required for normal SuS gene expression (Purcell et al., 1998). On the protein level, the enzyme activity in sucrose synthesis and sucrose cleavage was reported to be differentially affected by divalent metal ions (Delmer, 1972; Tsai, 1974; Huang and Wang, 1998), protein factors (Pontis and Salerno, 1982),
and the redox state of the enzyme (Pontis et al., 1981). In addition, SuS is post-translationally regulated by reversible phosphorylation. Phosphorylation of SuS in vivo and in vitro by endogenous or exogenous protein kinases in a Ca2+-dependent manner has been demonstrated in several plants such as maize (Huber et al., 1996; Winter et al., 1997; Subbaiah and Sachs, 2001), soybean nodule (Zhang and Chollet, 1997; Zhang et al., 1999), tomato (Anguenot et al., 1999), mung bean (Nakai et al., 1998), cotton (Haigler et al., 2001), and rice (Asano et al., 2002). Recently, Chikano et al. (2001) reported that an Arabidopsis SnRK3 protein, AtSR2, expressed in E. coli efficiently phosphorylated SuS in the presence of manganese ions. However, the significance of SuS phosphorylation in vivo is not clear. Phosphorylation/dephosphorylation of the enzyme has been suggested to play a role in regulating its activity (Zhang and Chollet, 1997; Nakai et al., 1998; Anguenot et al., 1999; Zhang et al., 1999; Haigler et al., 2001; Tanase et al., 2002), in its distribution between the cytosol, plasma membrane, and actin cytoskeleton (Amor et al., 1995; Winter et al., 1997; Winter and Huber, 2000), and also in the response of cells to environmental and developmental signals (Subbaiah and Sachs, 2001).
In rice, there are three non-allelic RSus genes encoding SuS (Wang et al., 1992; Yu et al., 1992; Huang et al., 1996; Wang et al., 1999). The gene products of RSus1 and RSus2 are ubiquitously present in suspension-cultured
*Corresponding author. Tel: +886-2-23630231 ext. 3251; Fax: +886-2-23660434; E-mail: aywang@ntu.edu.tw