Bot. Bull. Acad. Sin. (2003) 44: 211-216
He et al. Yeast application for controlling apple postharvest diseases
Yeast application for controlling apple postharvest diseases associated with Penicillium expansum1
Dan He, Xiao-Dong Zheng*, Yuan-Ming Yin, Ping Sun, and Hong-Yin Zhang
College of Biosystem Engineering and Food Science, Zhejiang University, 310029, Zhejiang, P.R. China
(Received August 23, 2002; Accepted May 8, 2003)
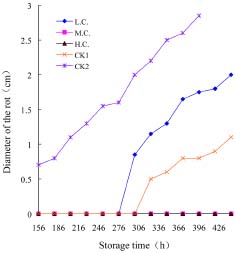
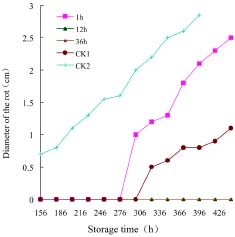
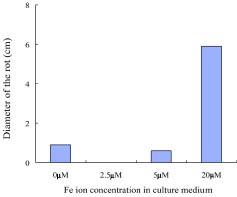
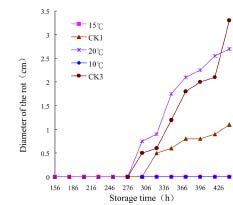
Abstract. Penicillium expansum is one of the main pathogens causing decay in fruits and vegetables. In recent years, researchers have discovered that some yeasts have antagonism against Penicillium expansum. In this study, two species of yeast that can biocontrol apple diseases caused by Penicillium expansum were discovered. At the same time, the antagonistic conditionssuch as yeast concentration, concentration of Fe2+ in the yeast culture medium, and inoculation timewere preliminarily studied, and the best conditions for yeast antagonism were determined.
Keywords: Apple; Biocontrol; Yeast.
Introduction
Apple is one of the most important fruits produced in China. To provide fruit throughout the year, fresh apples are stored after harvest. Postharvest losses caused by fungal diseases are the major factor limiting the storage life of apples. Postharvest fungal diseases of apple are mainly caused by Penicillium expansum (Romano et al., 1983). Traditionally, this disease is controlled by the application of synthetic fungicides (Eckert and Ogawa, 1988). However, the potential impact on environment as well as human health largely limits their application (Eckert et al., 1994). It is reported that some microbes become fungicide-resistant (Spotts and Cervantes, 1986; Holmes and Eckert, 1999), and thus a fungicide's effect on controlling fungal growth may be greatly reduced. Considering the human health and pollution risks, some fungicides are prohibited from use in many developed countries (e.g. America, England).
Recently, biological control has been developed as an alternative to synthetic fungicides (Wilson et al., 1993), and considerable success has been achieved by utilizing antagonistic microorganisms for controlling postharvest diseases. Ever since Guter reported that Bacillus subtilis was antagonistic toward fruit pathogens, many studies involving antagonistic microorganisms to control postharvest diseases of fruits and vegetables have been done (Wisniewski et al., 1991; Smilanick, 1992; Caztoria et al., 1997; Chand-Goyal and Spotts, 1997; Leibinger et al., 1997; Droby et al., 1998; Filnow, 1998; Benbow and Sugar,
1999; Sugar and Spotts, 1999; Ippolito and Franco, 2000). Roberts (1990) discovered that Cryptococcus laurentii has antagonistic activity against many postharvest pathogens. The competition for nutrients may play a role in the antagonism of Cryptococcus laurentii. Decay caused by Rhizopus sp. is reduced 70% when strawberries are treated with Aureobasiduim pullulans before storage (Lima et al., 1997). Calvente (Calvente et al., 1999) found that Rhodotorula glutinis produced rhodotorulic acid, which enhanced biocontrol activity of Rhodotorula glutinis against Penicillium expansum in postharvest apples. When applied to wounds 12 h after inoculation, Pseudomondia corragata significantly reduced brown rot in nectarines and peaches caused by Monilinia fructicola (Smilanick et al., 1993). Studies on postharvest biocontrol of fruits and vegetables have become an important new area of research.
Advantages of utilizing antagonistic microorganisms include reducing environmental pollutation, effectively controlling postharvest diseases, and producing high quality and safe food. Unfortunately, only a few studies in this field were reported in China (Fan and Tian, 2000a,b; Fan and Tian, 2001; Tian and Fan, 2001). There is a new opportunity to study biological control of postharvest diseases of fruits and vegetables in our country.
In this paper the capability of some kinds of yeast to control apple postharvest diseases caused by Penicillium expansum was studied, and the one with the best effect was identified. The antagonistic conditions of that yeast and the methods of antagonism were also discussed.
Materials and Methods
Materials
Fruit: Red Fushi Apple.
1Supported by Nation Natural Science Foundation of P.R. China (30170659).
*Corresponding author. Tel: 86-0571-86971167; Fax: 86-0571-86045315; E-mail: xdzheng@zju.edu.cn