Bot. Bull. Acad. Sin. (2003) 44: 217-222
Peng et al. Phylogenetic position of Dipentodon sinicus
Phylogenetic position of Dipentodon sinicus: evidence from DNA sequences of chloroplast rbcL, nuclear ribosomal 18S, and mitochondria matR genes
Yalin Peng1, Zhiduan Chen2, Xun Gong3, Yang Zhong4, and Suhua Shi1,*
1Key Laboratory of Gene Engineering of The Ministry of Education, Zhongshan University, Guangzhou 510275, The People's Republic of China
2 Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 100873, The People's Republic of China
3 Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650204, The People's Republic of China
4 Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, School of Life Sciences, Fudan University, Shanghai 200433, The People's Republic of China
(Received June 25, 2002; Accepted February 24, 2003)
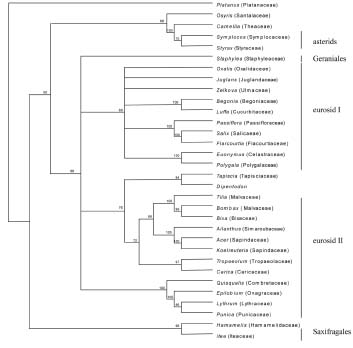
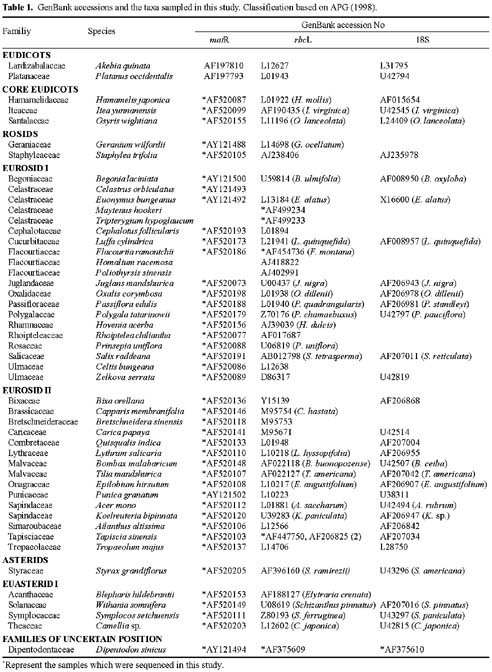
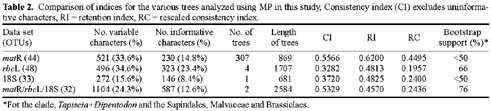
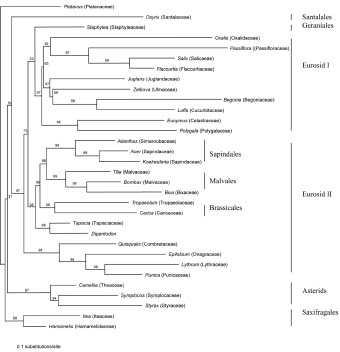
Abstract. Phylogenetic position of the monotypic genus Dipentodon has long been controversial. We investigated its position with 125 accessions representing 50 genera and 40 families of eudicots in the APG system. Four data setsincluding the chloroplast gene rbcL, the nuclear 18S ribosomal DNA, and the mitochondrial gene matR, as well as the combined matrixwere used in the study with the maximum parsimony (MP) and Bayesian inference (BI) analyses. The phylogenetic trees based on individual genes and the combined data suggested that Dipentodon is sister to Tapiscia (Tapisciaceae) and that Dipentodon could be placed in euroside II of the APG system. The clade of Dipentodon and Tapiscia is closest to Malvales and Sapindales. Such finding does not support the previously suggested close relationship between Dipentodon and various other groups, including Celastraceae, Samydaceae, Flacourtiaceae, Hamamelidaceae, and Santalales.
Keywords: Bayesian inference; Chloroplast rbcL; Dipentodon sinicus; Maximum parsimony; Mitochondrial matR; Molecular phylogeny; Nuclear ribosomal 18S.
Introduction
The genus Dipentodon Dunn consists of a single species D. sinicus Dunn, native to southern China and adjacent Burma and northeastern India (Merrill, 1941; Fischer, 1941; Li, 1986; Thorne, 1992; Bhattacharya and Johri, 1998). Its systematic position has been controversial since it was established and placed in the family Celastraceae in 1911 by S. T. Dunn. For example, Sprague (1925) moved it into the family Samydaceae based on the same variation range of floral base numbers in Dipentodon and Samydaceae. Many authors put the genus into the family Flacourtiaceae (including Samydaceae) (Fischer, 1941; Loesener, 1942; Metcalfe and Chalk, 1950; Lobreau, 1969). Record (1938) considered that Dipentodon is close to Hamamelidaceae based on the wood anatomic characters. Merrill (1941) proposed an independent family Dipentodontaceae Merr. and placed the family in Rosales between Hamamelidaceae and Rosaceae (Merrill, 1941;
Hutchinson, 1959, 1973; Schultze-Motel, 1964; Dahlgren, 1980; Cronquist, 1981; Takhtajan, 1987, 1997). However, Cronquist (1981) put the Dipentodontaceae into the order Santalales based on similar characters of the gynoecial structure. In the update Angiosperm Phylogeny Group (APG) classification scheme of flowering plants, the phylogenetic position of Dipentodontaceae is still uncertain (Angiosperm Phylogeny Group, 1998; APG II, 2003).
Recently, DNA sequences of the chloroplast, nuclear ribosomal, and mitochondrial genes have been widely used in plant phylogenetics, especially in reconstructing angiosperm phylogeny (Hoot et al., 1995; Qiu et al., 1999; Soltis et al., 1998; Kuzoff and Gasser, 2000). More importantly, combining multiple genes from three genomes in plants has proved effective in reducing homoplasy generated by gene-, function-, and genome-specific molecular evolutionary phenomena (Qiu et al., 1999). However, phylogenetic information about Dipentodon is poorly known from molecular data. In the present paper, therefore, we conduct phylogenetic analyses of the rbcL, 18S, and matR sequences to determine the phylogenetic position of Dipentodon and its relationships with related groups.
*Corresponding author. Tel: 86-20-84113677; Fax: 86-20-84113652; E-mail: lssssh@zsu.edu.cn