Bot. Bull. Acad. Sin. (2003) 44: 239-244
Qin et al. Phenological complementarity, species diversity, and ecosystem function
A reexamination of the relationships among phenological complementarity, species diversity, and ecosystem function
Guanglian Qin1,3, Guozhen Du1,*, Yanjiang Luo1, Gaosheng Dong2, and Jianyun Ma2
1State Key Laboratory of Arid Agroecology, Lanzhou University, Lanzhou 730000, Gansu Province, P.R. China
2Grassland Workgroup of Maqu County, Maqu 747000, Gansu Province, P.R. China
3College of Science, Huazhong Agriculture University, Wuhan 430070, Hubei Province, P.R. China
(Received September 3, 2002; Accepted March 4, 2003)
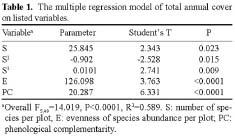
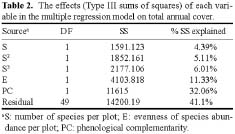
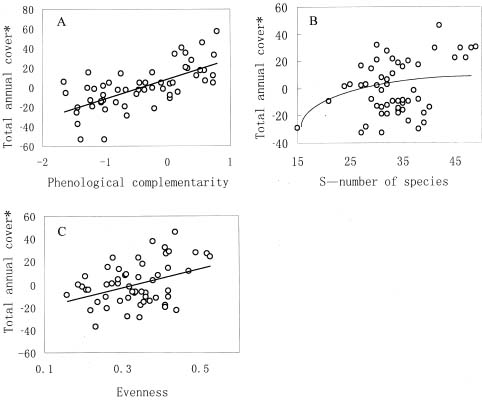
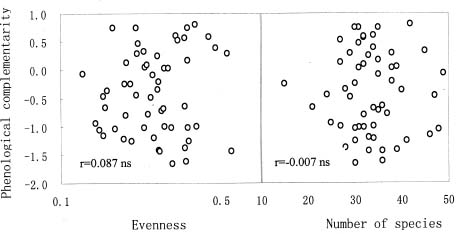
Abstract. Interest in the effects of species diversity on ecosystem function has been rapidly increasing, and many studies have concluded that increasing species diversity improves ecosystem function. Theories predict that phenological complementarity, the asynchrony of species resource use and growth, may explain how species diversity enhances ecosystem function but it remains largely untested. In this paper, we tried to determine: (i) whether, in a species-rich natural assemblage, increasing species diversity enhances ecosystem function by increasing phenological complementarity, and (ii) how phenological complementarity was related to the scales of alpha-diversity. Over a three-year period, we conducted the experiment in 55 permanent plots with a relatively homogeneous environment in natural alpine grasslands, and estimated percent cover of each plant species present in each plot three times in each growing season. Species diversity in each permanent plot was quantified in two ways, the total number of species, and the evenness of species abundance. Phenological complementarity was quantified as the negative logarithm of the variance ratio. Ecosystem function was measured as total annual cover (i.e. the sum of maximum cover values recorded for each species in a growing season). Statistical analysis of the relationships between total annual cover, species diversity, and phenological complementarity found that total annual cover increased with species diversity, and phenological complementarity was responsible for the positive effects of species diversity on total annual cover. Phenological complementarity, however, was not significantly related to species diversity. These results indicate that phenological complementarity was critical to the maintenance of total annual cover in our studied alpine plant communities. However, considering different mechanisms of species coexistence (e.g. local similarity), phenological complementarity is not necessarily linearly correlated with the scales of alpha-diversity in species-rich plant assemblages. In addition, other mechanisms, for example, the sampling effect for phenological complementarity, may simultaneously act on how phenological complementarity enhances ecosystem function. Therefore, greater understanding of how diversity and phenological complementarity influence ecosystem function will require more extensive experiments that control the effects of composition.
Keywords: Alpine grassland; Ecosystem function; Evenness; Phenological complementarity; Species diversity; Variance ratio.
Introduction
Recently, interest in the effects of species diversity on ecosystem function has been rapidly increasing, and researchers studying experimental terrestrial ecosystems (Naeem et al., 1995, 1996; Tilman et al., 1996, 1997a; Hector et al., 1999) have recently concluded that increasing species diversity improves ecosystem function. However, these studies have generated considerable criticism based upon their experimental methodology (Huston, 1997; Wardle, 1999), analysis (Aarssen, 1997), and general conclusions (Grime, 1998). As a result, understanding the mechanisms underlying the relationship between species diversity and ecosystem function remains important. Complementarity, where ecological differences (e.g., phenology) among species lead to more complete utiliza
tion of resources, may explain the biological effects of species diversity on ecosystem function (Tilman et al., 1997b; Loreau, 1998). Though theoretically attractive, few empirical diversity-ecosystem function studies have been constructed to test this mechanistic hypothesis (Hooper, 1998; Fridley, 2001).
Phenological complementarity, the asynchrony of species resource use and growth, may enhance ecosystem function when phenological differences allow different species to peak at different times throughout a growing season (Gulmon et al., 1983; Hooper, 1998). A previous empirical work showed that naturally occurring variation in species diversity had little effect on whether phenological complementarity can enhance ecosystem function (Stevens and Carson, 2001). This result emerged even though experimental manipulations of resources created contrasting environmental conditions analogous to larger scale environmental variation, thereby hopefully improving the generality of the findings (Stevens and Carson,
*Corresponding author. Tel: 086-0931-8912890; Fax: 086-0931-8912823; E-mail: Guozdu@Lzu.edu.cn