Bot. Bull. Acad. Sin. (2004) 45: 101-109
Su et al. Expression analyses of RP10
Expression analyses of a rice 10 kDa sulfur-rich prolamin gene
Pai-Hsiang Su1 , Su-May Yu2, and Ching-San Chen1,3,*
1Institute of Botany, Academia Sinica, Nankang, Taipei 115, Taiwan
2Institute of Molecular Biology, Academia Sinica, Nankang, Taipei 115, Taiwan
3Department of Agricultural Chemistry, National Taiwan University, Taipei 106, Taiwan
(Received October 15, 2003; Accepted January 6, 2004)
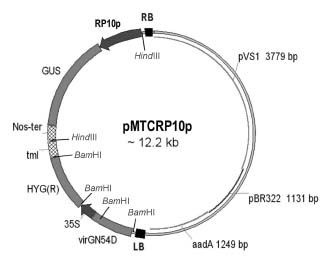
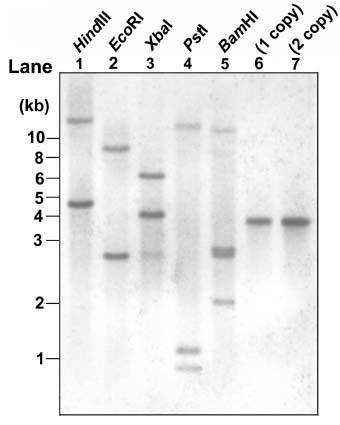
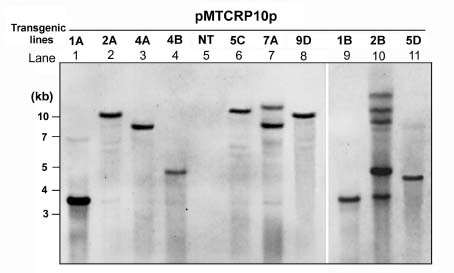
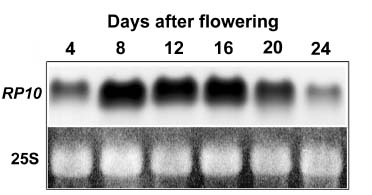
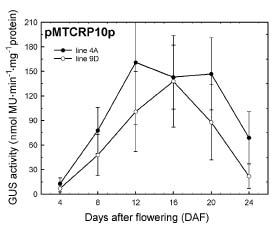
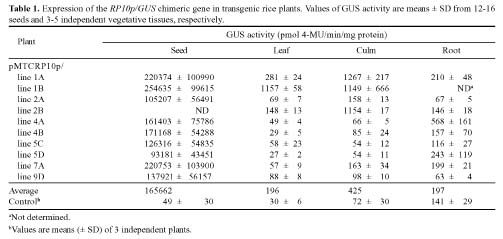
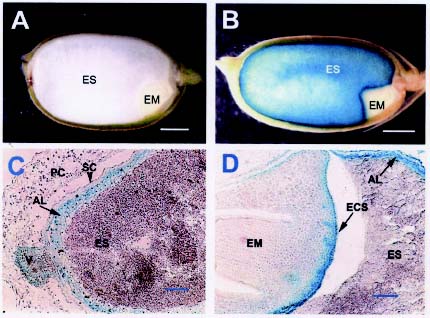
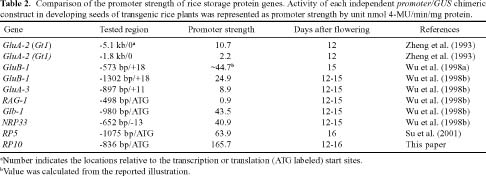
Abstract. Expression analyses of genes encoding sulfur-rich 10 kDa rice prolamins have not been reported to date although the entire genome of the model monocot plant has been sequenced in recent years. We isolated a 10 kDa rice prolamin gene, RP10, by genomic PCR, and the gene was registered in GenBank under the accession number AF294580. Genomic Southern-blot and bioinformatic study revealed that genes encoding RP10 homologes (class IV rice prolamins) were composed of two to three copies per haploid genome in Japonica cultivars Tainung 67 and Nipponbare. To elucidate the temporal and spatial expression of RP10, we introduced a chimeric gene that consisted of the 836 bp upstream sequence of RP10 and the coding regions of b-D-glucuronidase (GUS) into rice via Agrobacterium tumefaciens mediated transformation. The expression levels of GUS followed the accumulation pattern of endogenous RP10 transcript in parallel, indicating that the expression of GUS did reflect the endogenous temporal control of RP10 gene. Maximal GUS activity was reached at 12~20 days after flowering (DAF) in maturing seeds. Histochemical analysis showed that the specific expression of GUS in seeds was not restricted to the endosperm cells, but also occurred in the vascular bundle and epithelial cells of scutellum. In comparison with promoters of other rice storage protein genes, RP10 promoter exhibited a high expression level, with a long plateau period. Our studies suggest that RP10 promoter could be potentially useful for over-expression of foreign genes in transgenic rice seeds.
Keywords: Epithelial cells of scutellum; Oryza sativa; Prolamin RP10; Promoter; Seed-specificity; Storage proteins; Transformation.
Abbreviations: DAF, days after flowering; GUS, b-glucuronidase; 4-MU, 4-methylumbelliferone; PB-I, protein body I; X-Gluc, 5-bromo-4-chloro-3-indolyl-b-D-glucuronide.
Introduction
Prolamin is one of the major seed storage proteins in rice grains (Krishnan and White, 1995). It consists of three polypeptide groups, 10, 13, and 16 kDa, which all accumulate in the protein body I (PB-I) localized in the starchy endosperm. Each group has further micro-heterogeneity resulting from the corresponding gene subfamily (Tanaka et al., 1980; Ogawa et al., 1987). Analysis of amino acid composition revealed that the 13 kDa prolamin peptides, the major group, are rich in glutamic acid, glutamine, and leucine, but are poor in methionine, cysteine, and lysine. In contrast to the 13 kDa prolamin, the 10 and 16 kDa prolamins are rich in sulfur-containing amino acids. The 10 kDa prolamin, in particular, contains a large amount of methionine and cysteine (Hibino et al., 1989).
Ultrastructural analysis by electron microscopy shows that PB-I is a spherical concentric ring structure with a dense center (Bechtel and Juliano, 1980; Ogawa et al., 1987). In light of the staining mechanism, the dense center and dotted area were suggested to be the deposition
site of reducing substances, possibly contributed by 10 and/or 16 kDa sulfur-rich prolamins (Hibino et al., 1989). Research on the digestibility of rice grain revealed that the dense center of PB-I was resistant to pepsin treatment and indigestible by the human digestive tract (Bechtel and Pomeranz, 1978; Tanaka et al., 1978). Therefore, the isolation and characterization of the 10 kDa prolamin polypeptides and genes are important for understanding the formation of PB-I and genetic engineering to improve the digestibility of rice grain.
The cDNA clone lRP10, isolated from the japonica rice cultivar Nipponbare and encoding the 10 kDa sulfur-rich prolamin, was first reported by Masumura et al. (1989). A polypeptide sequence composed of 134 amino acids was deduced from the nucleotide sequence. After removing the signal peptide, the mature polypeptide contains 20% methionine and 10% cysteine (Masumura et al., 1989). Thereafter, several genes encoding the 10 kDa sulfur-rich prolamin peptides were cloned from different rice cultivars (Masumura et al., 1989; Feng et al., 1990; Wang et al., 1994; You et al., 1995). Comparison of these cloned genes revealed that nucleotide and deduced amino acid sequences share more than 95% and 93% homology, respectively. Based on the unique amino acid composition, which is rich in sulfur-containing amino acids, some workers have at
*Corresponding author. Tel: +886-2-27899590 ext. 251; Fax: +886-2-27858936; E-mail: chingsan@gate.sinica.edu.tw