Bot. Bull. Acad. Sin. (2004) 45: 143-147
Chou and Chang _ Seed germination of orchids
Asymbiotic and symbiotic seed germination of Anoectochilus formosanus and Haemaria discolor and their F1 hybrids
Ling-Chin Chou and Doris Chi-Ning Chang*
Department of Horticulture, National Taiwan University, Taipei, Taiwan 10617, ROC
(Received July 15, 2003; Accepted February 23, 2004)
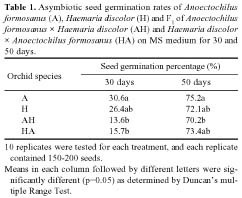
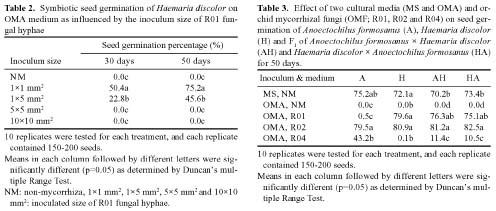
Abstract. Two orchid species Anoectochilus formosanus Hayata and Haemaria discolor var. dawsoniana, which have different flowering times in nature, were induced to flowering synchronously by controlling the culture temperature and light duration in the phytotrons. They were crossed, and the F1 hybrid seeds were collected and named as H. discolor × A. formosanus (HA) and A. formosanus × H. discolor (AH). It was found that fifty days after the asymbiotic culture in MS medium, the germination rates of all the four kinds of seed were higher than 70%. For the symbiotic seed germination on oatmeal agar (OMA) medium, only the R02, as compared to the R01 and R04 isolates of Rhizoctonia spp., had a more than 80% germination rate. SEM observation of various seed germination stages for the non-mycorrhizal and the mycorrhizal protocorms of A. formosanus showed that 70 days after seed sowing, the inoculated seeds developed to a more advanced stage than the non-inoculated control. Asymbiotic and symbiotic germination occurred after the uptake of water, and the seed coat was ruptured by the enlargement of embryo. Afterward the papilla, protocorm and apical meristem appeared. In the symbiotic germinated embryos, the fungal hyphae penetrated the protocorm and formed pelotons, which was the tolypophagy type of infection. No hypha infection was found in the asymbiotic germinated embryos.
Keywords: In vitro propagation; Orchid mycorrhizal fungi; Orchidaceae; Jewel orchid; Scanning electron microscopy; Seed germination.
Introduction
Both Haemaria discolor var. dawsoniana and Anoectochilus formosanus Hayata belong to orchidaceae and were perennial and medicinal herbs. The whole H. discolor plant has medicinal value in lung protection, and suffers little from disease or insect problems. It is also more heat resistant than A. formosanus, which is highly susceptible to diseases and is non-heat resistant. Anoectochilus formosanus is regarded as " the king of medicines " by aborigines in Taiwan because of its diverse pharmacological effects, such as liver protection, cancer prevention, blood sugar reducing for diabetes, and the treatment of cardiovascular diseases etc. (Kan, 1986; Shiau et al., 2002). Haemaria discolor blooms from late December to March (Chou, 1997) while A. formosanus generally blooms from August to November. In order to enhance the heat-tolerance and disease resistance of A. formosanus, the breeding of these two species of orchid plants was conducted. We hope that an F1 hybrid of H. discolor × A. formosanus and A. formosanus × H. discolor will be heat-tolerant and yet still maintainits medicinal value. Although F1 hybrids of Anoectochilus sikkimensis × Ludisia discolor have been reported (Belitsky and Bersenev, 1999), there are no reports concerning the symbiotic germination of the F1 hybrid seeds. In our previous studies, we have shown that
these two orchids could be propagated from seeds through symbiotic and asymbiotic germinations (Chou, 1997). Besides, we have found that the growth of these two orchids was stimulated by Rhizoctonia spp. (Chou, 1997; Tsai, 1997). The present study reports a feasible and effective method for the germination of A. formosanus, H. discolor and their F1 hybrid seeds through asymbiotic and symbiotic cultures. The asymbiotic and symbiotic germination processes are also investigated by scanning electron microscopy (SEM).
Materials and Methods
Plant Materials and Culture Conditions
Plants of H. discolor and A. formosanus were cultivated for eight months under 25/20°C day/night temperature in the phytotron. Then they were put into the phytotron at 23/18°C and a 16-h photoperiod of 3,000-4,000 Lux fluorescent light in mid May to induce blooming. In late October, both kinds of plants could flower synchronously. Between these two orchids, hand pollination was conducted. After pollination for 40 days, capsules of H. discolor (H), A. formosanus (A), H. discolor × A. formosanus (HA) and A. formosanus × H. discolor (AH) were collected. Capsules were surface-sterilized with a cotton ball swab moistened with 70% ethanol, followed by disinfesting under agitation for 15 min in 1% sodium hypochlorite, and were rinsed three times with sterile distilled water. About 200-300 seeds from disinfested capsules
*Corresponding author. Tel. & Fax: +886-2-23690482; E-mail:hyha@ccms.ntu.edu.tw