Bot. Bull. Acad. Sin. (2004) 45: 187-196
Hsu et al. Rice starch phosphorylase purification and characterization
Purification and characterization of a cytosolic starch phosphorylase from etiolated rice seedlings
Jen-Hung HSU, Chien-Chih YANG, Jong-Ching SU, and Ping-Du LEE*
Biochemistry Laboratory, Department of Agricultural Chemistry, and Department of Biochemical Science and Technology, National Taiwan University, No.1 Roosevelt Rd., Sec 4, Taipei 106, Taiwan
(Received April 28, 2003; Accepted May 18, 2004)
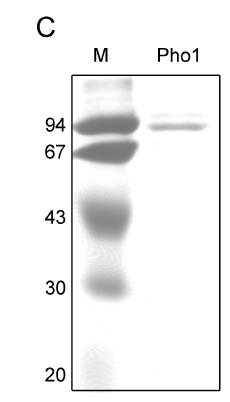
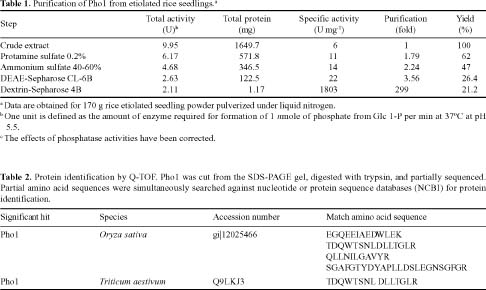
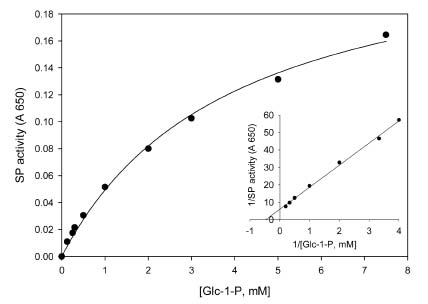
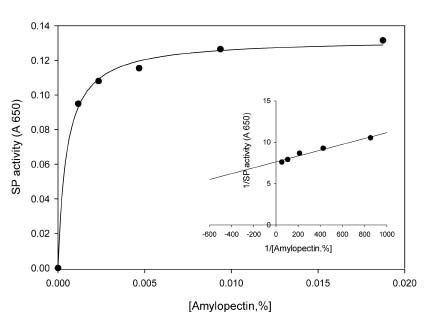
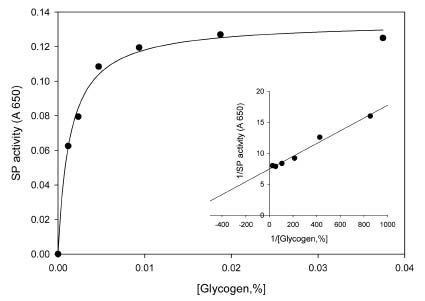
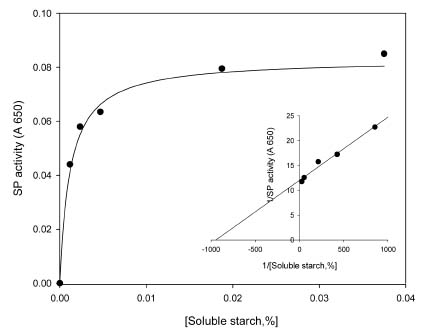
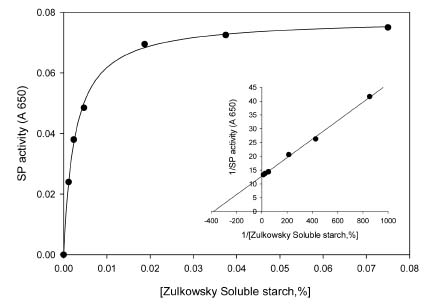
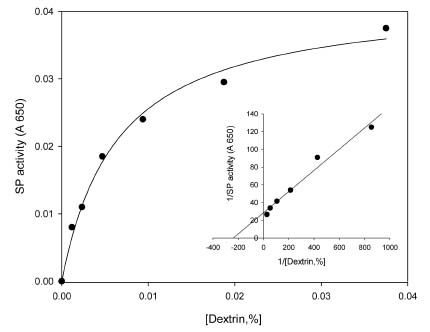
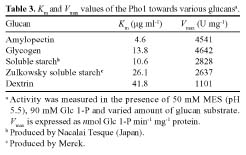
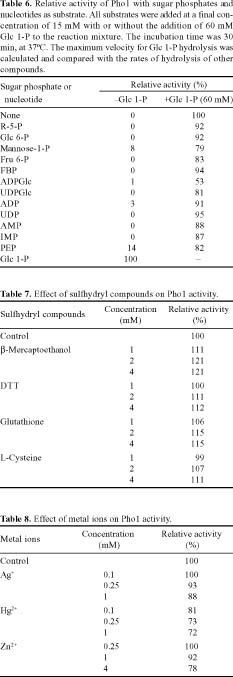
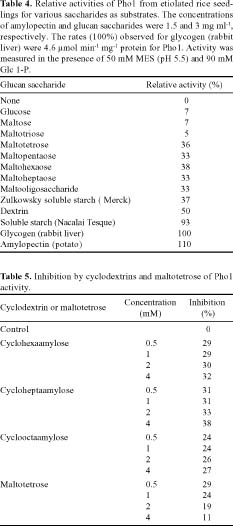
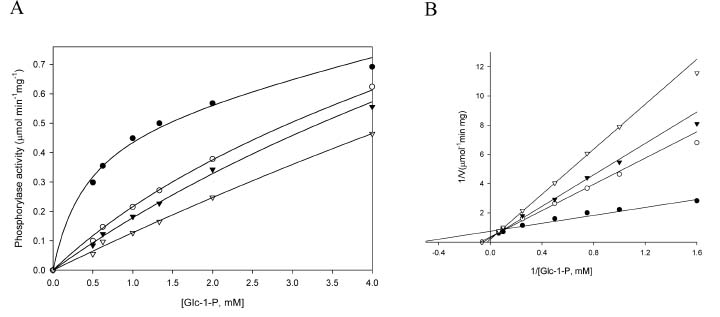
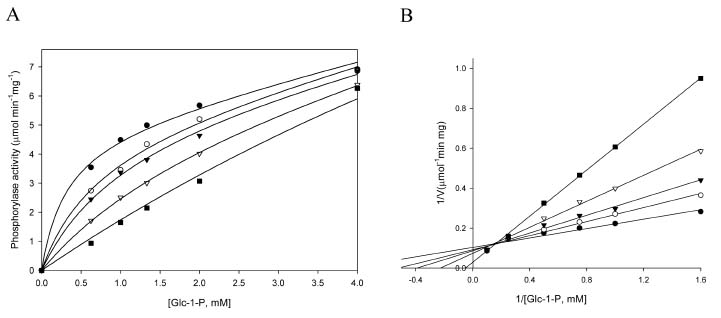
Abstract. Starch phosphorylase1 (Pho1) from etiolated rice (Oryza sativa L. cv. Tainong 67) seedlings was purified by ammonium sulfate fractionation, DEAE-Sepharose CL-6B anion exchange chromatography, and dextrin-Sepharose 4B affinity chromatography. The purification fold was 299, and the enzyme activity recovery was about 21%. The molecular mass of the native Pho1 on Superose 12 gel filtration was 145 kDa. The subunit molecular weight as determined by SDS-PAGE was 85 kDa. The enzyme has an optimum pH of 5 and an optimum reaction temperature of about 45oC~50 oC. In the synthetic reaction for Glc 1-P, the Km value was 2.1 mM, and the Vmax value was 5.85 U mg-1. In the phosphorolytic direction for orthophosphate, the Km value was 3.8 mM. Pho1 has a higher affinity for amylopectin, glycogen, soluble starch and dextrin than for maltooligosaccharide (6 to 10 glucose units). In addition, the Km value for amylopectin was ninefold lower than for dextrin. Cyclohexaamylose, cycloheptaamylose, cyclooctaamylose, and maltotetrose were inhibitors of Pho1. Mannose 1-P, Fru 6-P, ADPGlc, UDPGlc, AMP, IMP and PEP also inhibit Pho1. The metal ions Ag+, Hg2+ and Zn2+ also reduce the enzyme activity. However, thiol reagents activate Pho1 activity, suggesting that sulfhydryl-group(s) may be required for enzyme stability.
Keywords: Affinity chromatography; Dextrin; Glucan; Rice; Starch phosphorylase.
Abbreviations: Pho1, H type starch phosphorylase; Pho2, L type starch phosphorylase; Glc 1-P, glucose 1-phosphate.
Introduction
The plant starch phosphorylases (Pho, SP, a-glucan phosphorylase, EC 2.4.1.1) catalyze the reversible phosphorolysis of a-1,4-glucans and have been reported in many higher plants (Fukui, 1983; Steup and Schachtele, 1986). The reaction can be described essentially by the following equation:
(a-1,4-D-glucose)n + Glc 1-P (a-1,4-D-glucose)n+1 + Pi
A glucosyl unit is transferred from Glc 1-P to starch of increasing chain length, and this is termed as "the synthetic direction." In the other direction, the addition of inorganic phosphate generates Glc 1-P from starch as the chain length decreases, and this is called "the phosphorolytic direction."
Pho is generally regarded as a starch-degrading enzyme, but
some evidence suggests it might also have an important synthetic function. In
most plants, Pho has been found in both cytosolic and amyloplastic forms in
pea seeds and leaves (Hanes, 1940), broad beans (Suda et al., 1987), potato
tubers (Hanes, 1940), maize (Tsai and Nelson, 1968; Mu et al., 2001), barley
(Baxter and Duffus, 1973), rice seeds (Richardson and Matheson, 1977), spinach
(Steup and Latzko, 1979), sweet potato (Chang et al., 1987; Lu et al., 1995),
bananas (Richardson and Matheson, 1977; Da Mota ![]()
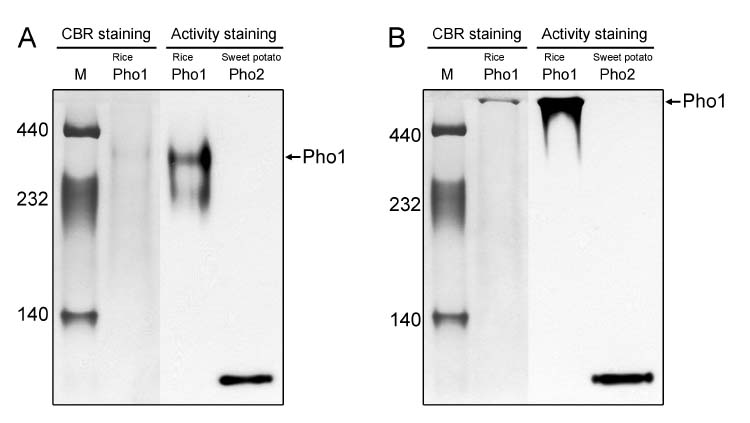
et al., 2002), and seaweed (Fredrick, 1973; Yu and Pedersen, 1991). Specifically, the Pho1 or H (high affinity for glycogen) type of starch phosphorylase that occurs in the cytosol of plant cells has a molecular mass of about 90 kDa. Pho2 or L (low affinity for glycogen) form, is located in the amyloplast and has a molecular mass of about 100 kDa (Steup and Latzko, 1979; Conrads et al., 1986; Mu et al., 2001; Da Mota et al., 2002).
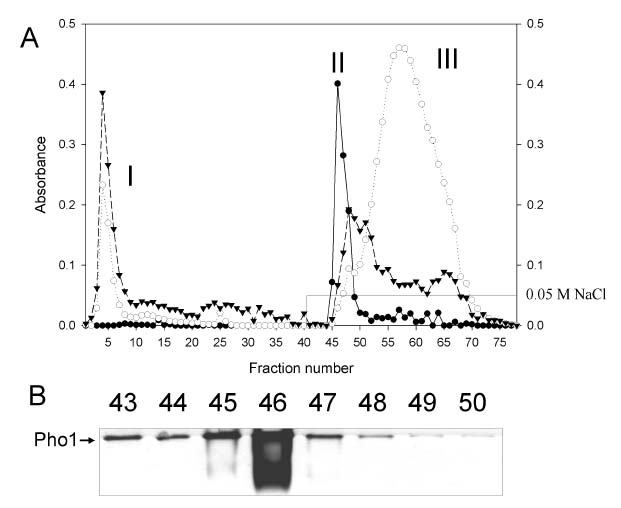
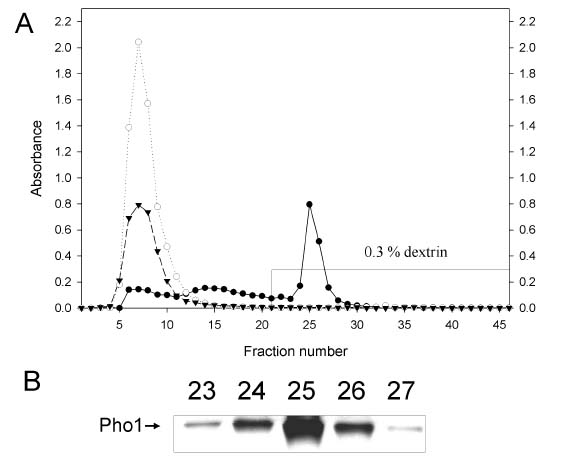
Here we report the purification and enzymatic characterization of a rice starch phosphorylase, Pho1. Since the level of Pho1 protein in etiolated rice seedlings is very low, we developed a procedure that greatly increases the yield and results in a Pho1 enzyme of high purity. Pho1 purified by this procedure was analyzed, and its molecular mass and kinetic properties was determined.
Material and Methods
Materials
Rice seeds (Oryza sativa L. cv. Tainong 67) were grown in the dark for 14 days at 30oC in a growth chamber. These fresh etiolated rice seedlings were used for enzyme purification. Imidazole, protease inhibitors, protamine, MES, glucan saccharides, cyclodextrins, sugar phosphate, nucleotides, thiol reagents, and ammonium molybdate were obtained from Sigma (USA). Soluble starch was purchased from Nacalai Tesque Inc. (Kyoto, Japan). Zulkowsky soluble starch, AgNO3, ammonium sulfate, Tris-Cl were supplied
*Corresponding author. Tel:+886-2-2363-3610; Fax: +886-2- 2363-3610; E-mail: pingdu@ntu.edu.tw