Bot. Bull. Acad. Sin. (2004) 45: 291-299
Kuo and Kao Antioxidant enzymes and Cd toxicity in rice seedling
Antioxidant enzyme activities are upregulated in response to cadmium in sensitive, but not in tolerant, rice (Oryza sativa L.) seedlings
Mei Chun KUO and Ching Huei KAO*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received January 14, 2004; Accepted May 3, 2004)
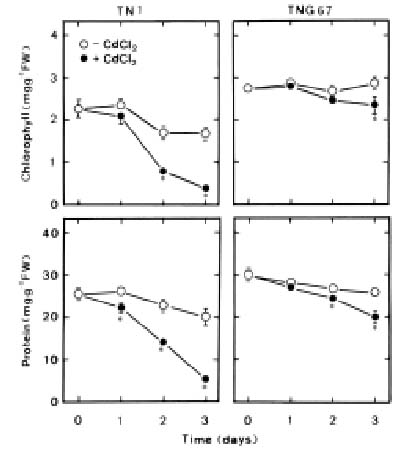
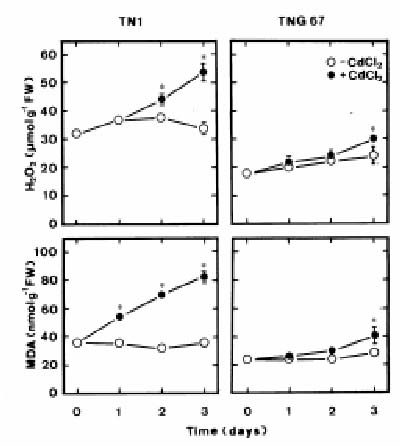
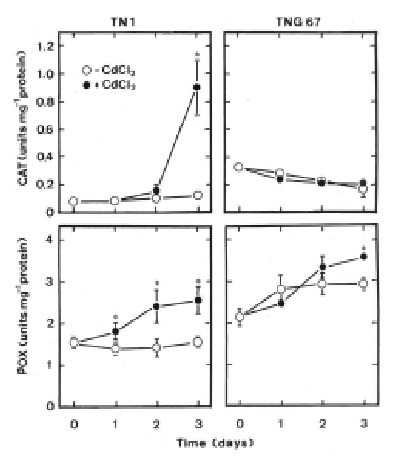
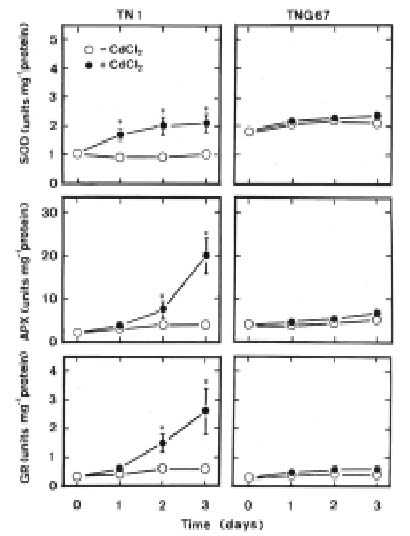
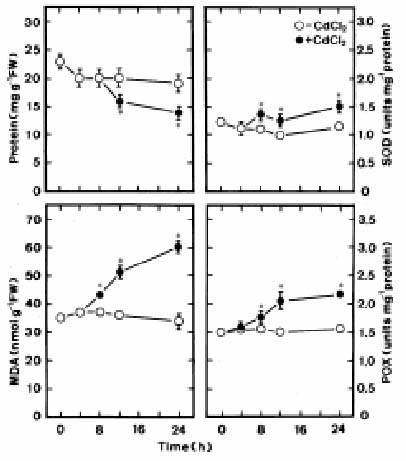
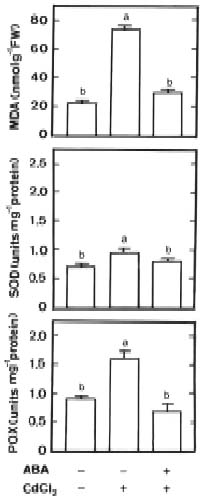
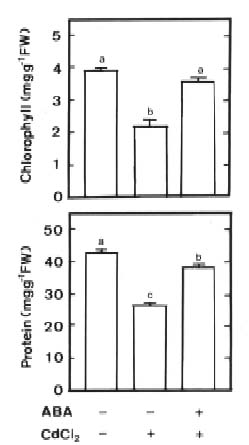
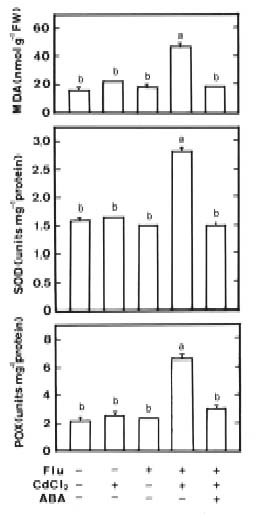
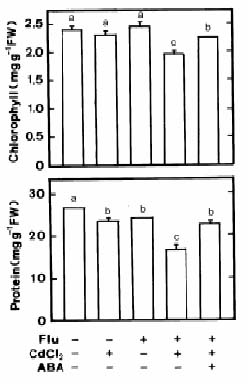
Abstract. Changes in H2O2 and malondialdehyde (MDA) contents and antioxidant enzyme activities in Cd-treated rice (Oryza sativa L.) seedlings of two cultivars were investigated. On treatment with CdCl2, increases in H2O2 and MDA contents and antioxidant enzyme activities [speroxide dismutase (SOD), ascorbate peroxidase, glutathione reductase, catalase, and peroxidase (POX)] were observed in the leaves of Cd-sensitive cultivar (cv. Taichung Native 1, TN1) but not in Cd-tolerant cultivar (cv. Tainung 67, TNG67). The increased content of MDA and activities of SOD and POX preceded the occurrence of toxicity in CdCl2-treated TN1 leaves. Pretreatment with abscisic acid (ABA) enhanced Cd tolerance and reduced Cd-induced increase in the content of MDA and increase in the activities of SOD and POX in TN1 leaves. Exogenous application of ABA biosynthesis inhibitor, fluridone, decreased Cd tolerance, increased the content of MDA, and increased the activities of SOD and POX in Cd-treated TNG67 leaves. Furthermore, fluridone's effects on toxicity, the content of MDA, and the activities of SOD and POX in Cd-treated TNG67 leaves were reversed by the application of ABA. In conclusion, the oxidative stress is differently expressed in TN1 and TNG67 rice seedlings in response to CdCl2. Results also suggest that CdCl2 causes an oxidative stress and CdCl2-induced toxicity is mediated through oxidative stress in TN1 leaves.
Keywords: Abscisic acid; Cadmium; Oryza sativa L.; Oxidative stress.
Abbreviations: ABA, abscisic acid; APX, ascorbate peroxidase; CAT, catalase; Flu, fluridone; FW, fresh weight; GR, glutathione reductase; MDA, malondialdehyde; POX, peroxidase; SOD, superoxide dismutase; TN1, Taichung Native 1; TNG67, Tainung 67.
Introduction
Oxygen is essential for the existence of aerobic life, but toxic active oxygen species (AOS), which include the superoxide anion (O2-), hydroxyl radical (OH.) and hydrogen peroxide (H2O2), are generated in all aerobic cells during metabolic processes (Foyer et al., 1994; Asada, 1999). Injury caused by these AOS, known as oxidative stress, is one of the major damaging factors in plants exposed to environmental stress. Plants cope with oxidative stress by using antioxidant enzymes such as superoxide dismutase (SOD), ascorbate peroxidase (APX), glutathione reductase (GR), peroxidase (POX), catalase (CAT), and the low molecular weight antioxidants, ascorbic acid and glutathione (Noctor and Foyer, 1998; Asada, 1999).
Cadmium (Cd), a heavy metal toxic to humans, animals, and plants is a widespread pollutant with a long biological half-life (Wagner, 1993). Three lines of evidence indicate that one mechanism of Cd toxicity is related to oxidative
stress in plant cells. First, Cd can promote the generation of AOS (Piqueras et al., 1999; Romero-Puertas et al., 1999; Chen et al., 2000; Shah et al., 2001; Sandalio et al., 2001; Schützendübel et al., 2001; Olmos et al., 2003). Second, Cd can inhibit or stimulate the activities of antioxidant enzymes (Shaw, 1995; Gallego et al., 1996; Chaoui et al., 1997; Dixit et al., 2001; Shah et al., 2001; Iannelli et al., 2002; León et al., 2002). Third, treatment with Cd results in cellular oxidative damage or lipid peroxidation (Shaw, 1995; Gallego et al., 1996; Chaoui et al., 1997; Lozano-Rodríguez et al., 1997; Dixit et al., 2001; Shah et al., 2001; Chien et al., 2002).
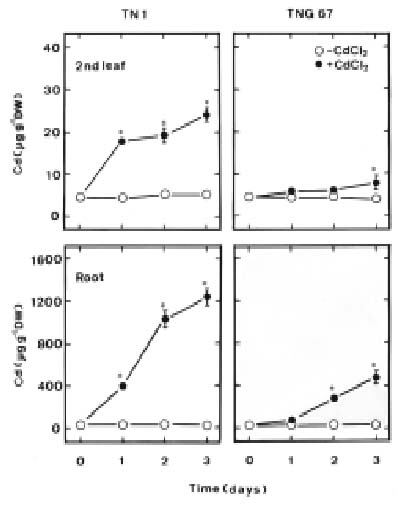
This study examines H2O2 content, lipid peroxidation, and antioxidant enzyme activities in two cultivars of rice (Oryza sativa L.) seedlings in response to Cd. One cultivar (cv. Taichung Native 1, TN1) is known to be sensitive to Cd, and the other (cv. Tainung 67, TNG67) is known to show significant tolerance to Cd (Hsu and Kao, 2003a). If a mechanism related to oxidative stress is involved in Cd toxicity, this mechanism should then be differently expressed in plants tolerant and sensitive to Cd. Therefore, the aim of this study was to investigate whether the oxidative stress mechanism is differently expressed in TN1 and TNG67 rice seedlings in response to Cd.
*Corresponding author. Tel: 886-2-23698159; Fax: 886-2-23620879; E-mail: kaoch@ntu.edu.tw