Bot. Bull. Acad. Sin. (2004) 45: 339-346
Cheng Statistical approaches on discriminating spatial variation of species diversity
Statistical approaches on discriminating spatial variation of species diversity
Chi-Chuan CHENG*
Division of Forest Management, Forestry Research Institute, Council of Agriculture, Taipei 100, Taiwan, Republic of China
(Received May 12, 2003; Accepted May 19, 2004)
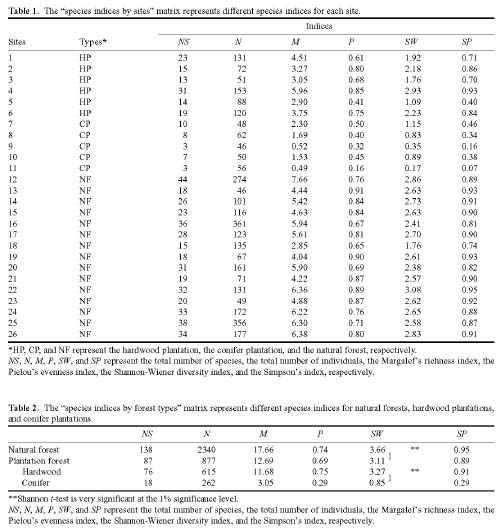
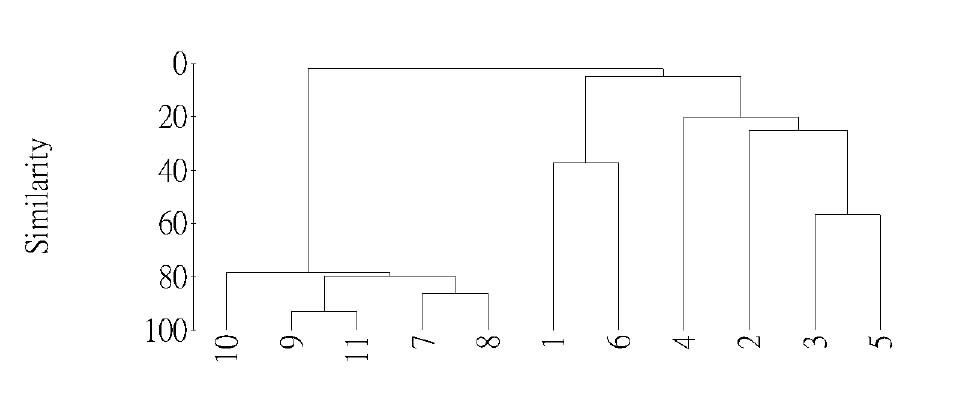
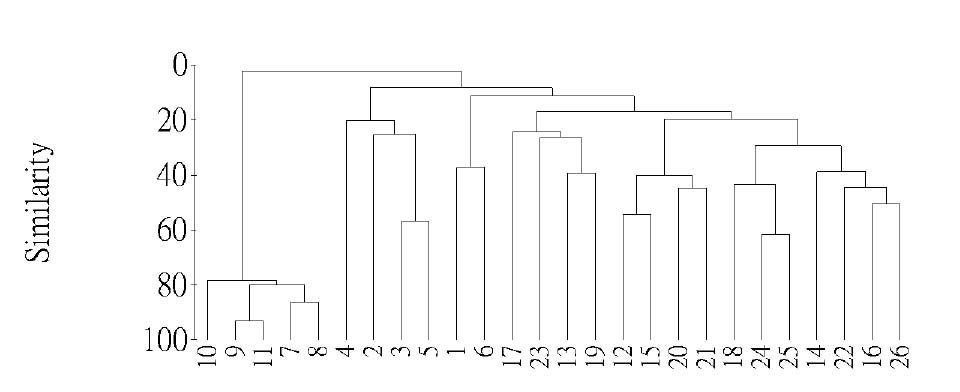
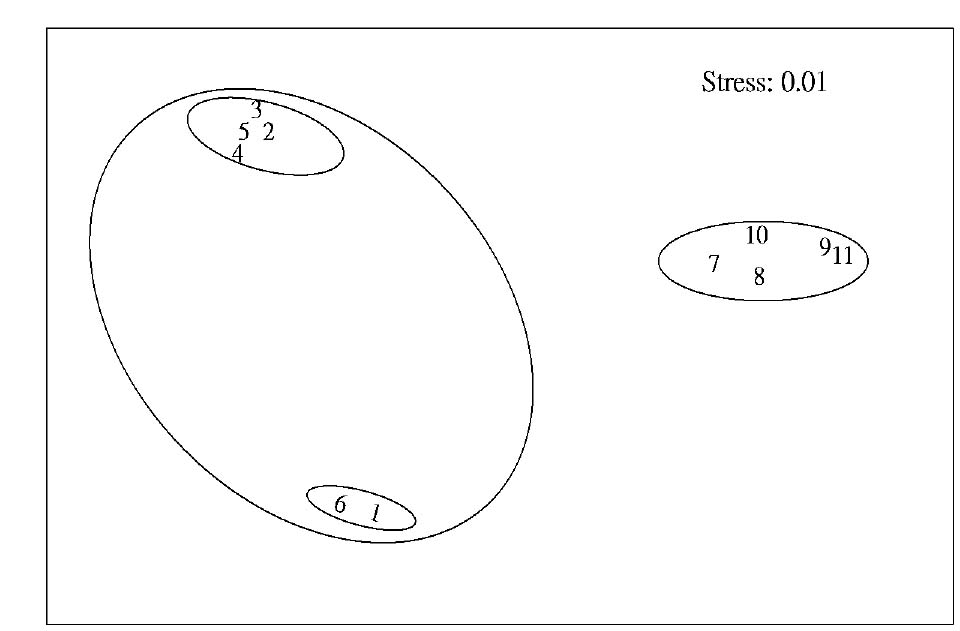
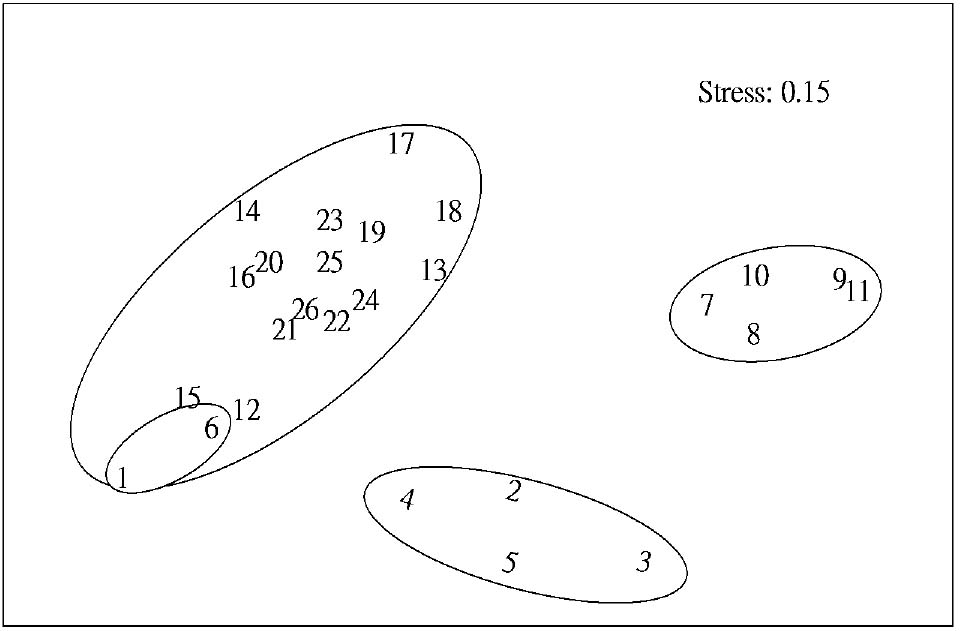
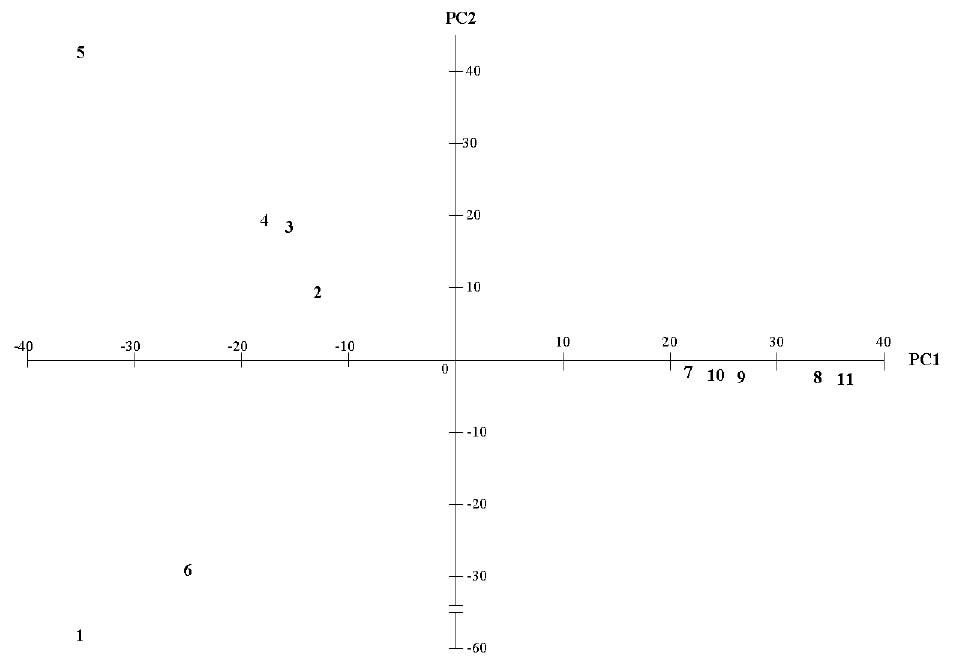
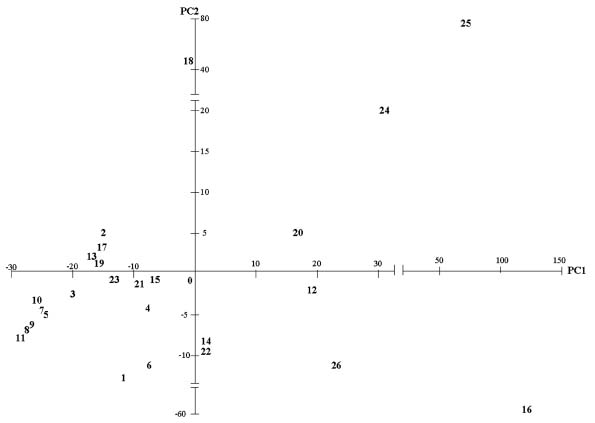
Abstract. This study applied statistical approaches to the discrimination of spatial variations between sites and between forest types in the upper area of the Liukuei Experimental Forest of Taiwan Forestry Research Institute, Taiwan. The main purpose was to compare the effectiveness of various statistical approaches and then present the best strategy for discriminating the spatial variations of species diversity. The two methods used were (1) univariate methods by diversity measures, Shannon t-test, and (2) multivariate methods by cluster analysis, ordination by non-metric multi-dimensional scaling, and principal component analysis. The results by univariate methods indicate that diversity differences exist between sites and between forest types. Meanwhile, the natural forest has more diversity than the plantation, and the hardwood plantation has more diversity than the conifer plantation. The differences between forest types are very significant at the 1% significance level according to the Shannon t-test. The results indicate that univariate methods by diversity measures are a flexible way to reduce the complexity of "species by sites" matrices into a single coefficient. The results of using multivariate methods indicate that cluster analysis and ordination by non-metric multi-dimensional scaling and principal component analysis are useful techniques for discriminating spatial variations. However, ordination by non-metric multi-dimensional scaling discriminates better than principal component analysis. In addition, ordination by non-metric multi-dimensional scaling is a more informative summary than cluster analysis, and the combination of both the analyses is more effective than either alone for the mutual consistency of representations. It is concluded that the most powerful tools for discriminating the spatial variations of species diversity are in the multivariate category. Among multivariate methods, ordination by non-metric multi-dimensional scaling is preferable, and its superimposition with cluster analysis is recommended in order to obtain more information regarding the relationship between sites and between forest types.
Keywords: Spatial variation; Species diversity; Statistical approach.
Introduction
Ecological research on the relationship between biodiversity and forest management practices has been increasingly emphasized (Burton et al., 1992; Halpern and Spies, 1995; Roberts and Gilliam, 1995). As for biodiversity, a variety of different indices (e.g., total number of individual, total number of species) can be used as measures of some attribute of community structure because they are often seen as ecological indicators (Magurran, 1988). However, these indices tend to be less informative and less amenable to simple statistical analysis than species diversity (Clarke and Warwick, 1994). Two different aspects of community structure contribute to species diversity, i.e., species richness and species evenness. Different species diversity indices emphasize species richness or evenness to varying degrees. Hill (1973) pointed out that several of these indices are used as special cases, and a good account of their relative merits and disadvantages
can be found in Magurran (1988). Among the species diversity indices, the Shannon-Wiener diversity index is the most commonly used because it incorporates both species richness and evenness components and can provide heterogeneity information for vegetation and wildlife studies (Rosenstock, 1998; Blair, 1999; Cheng, 1999). Also, it is possible to test the differences between two communities using a Shannon t-test (Hustcheson, 1970; Magurran, 1988; Cheng, 1999).
In most community structure studies, the "species-by-samples" matrices are typically large because an abundance of readings for a set of species is usually taken at a number of sites at one time (spatial analysis) and at the same site at a number of times (temporal analysis). For the data interpretation on community structure, several sophisticated statistical techniques for handling, interpreting, and analyzing "species-by-samples" matrices are used, ranging from the reduction of multi-dimensional data to simple diversity indices through distributional representations of richness, dominance, evenness, to multivariate approaches involving cluster analysis or ordination methods by multi-dimensional scaling (MDS) or principal component analysis (PCA).
*Corresponding author. Tel: 886-2-23039978 ext. 1318; Fax: 886-2-23754216; E-mail: cccheng@serv.tfri.gov.tw