Bot. Bull. Acad. Sin. (2005) 46: 35-42
SUNG et al. Capsaicin biosynthesis in water-stressed hot pepper fruits
Capsaicin biosynthesis in water-stressed hot pepper fruits
Yu SUNG*, Yu-Yun CHANG, and Ni-Lun TING
Department of Horticulture, National Chung-Hsing University, Taichung, Taiwan
(Received March 23, 2004; Accepted August 11, 2004)
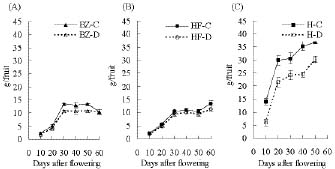
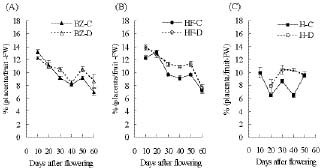
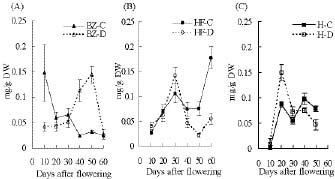
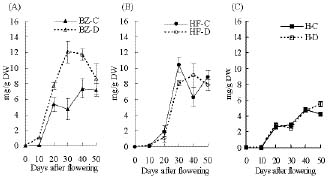
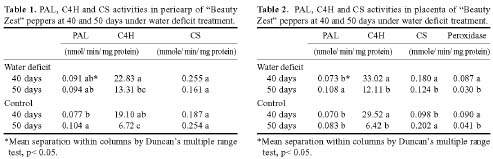
Abstract. "Hungariana," "Beauty Zest," and "Home Flavor" hot pepper plants (Capsicum annuum L. var. annuum) were grown with an ample or a limited water supply. The fruits of plants in the water deficit treatment were small, had a proportionally heavier placenta and had a higher concentration of capsaicin. The concentration of capsaicin in the placenta of "Beauty Zest" fruits in the water deficit treatment began to increase rapidly 10 days after flowering (DAF). It reached a maximum 30 days DAF and was 3.84-fold higher than in the placenta of control treatment plants. In the pericarp, the concentration of capsaicin reached a maximum 50 days DAF and was 4.52-fold higher than in the control treatment. In "Hungariana" fruits, the concentration of capsaicin in the placenta was not significantly different among treatments. Phenylalanine ammonia-lyase (PAL) activity was higher in the placenta of "Beauty Zest" fruits in the water deficit treatment than in the fruits of control plants 50 DAF. In 40 or 50 DAF, cinnamic acid-4-hydroxylase (C4H) activity was higher in plants subjected to the water deficit treatment than in control plants. In both treatments, C4H activity in placenta was 1.4 to 1.5-fold greater than in the pericarp 40 DAF. Capsaicinoid synthetase (CS) activity 40 DAF was 1.45 to 1.58-fold higher in fruits in the water deficit treatment than in fruits in the control treatment. Although peroxidase activity was lower in plants in the water deficit treatment than in the control treatment, the difference was not significant.
Keywords: Capsaicin; Hot pepper; Pungency; Water treatment.
Introduction
Hot pepper plants are shallow-rooted and lack a taproot, but they develop many fibrous roots that grow to 30 cm below the soil surface. Pepper plants are notorious for their sensitivity to moisture stress at flowering and fruit setting. The blossoms and immature pods of water-stressed plants often drop. On the other hand, water-stressed plants typically produce more pungent pods (Bosland and Votava, 2000).
Bucholz (1816) first discovered and extracted the pungent, oily substance from hot peppers. The active ingredient in hot pepper was isolated by Thresh (1846), and was named it capsaicin (Govindarajan, 1987). Today, more than 15 capsaicinoids are known. Capsaicin and dihydrocapsaicin account for more than 80% of the capsaicinoids that determine the pungency of peppers (Bernal et al., 1993b; Walpole at al., 1996; Kobata et al., 1998).
Two pathways are involved in the biosynthesis of capsaicinoids: fatty acid metabolism and the phenylpropanoid pathway (Ochoa-Alejo and Gomez-Peralta, 1993). The phenolic structure comes from the phenylpropanoid pathway, in which phenylalanine is the precursor. The formation of ferulic acid from phenylalanine is well understood in other higher plants. Four enzymes, phenylalanine ammonia-lyase (PAL), cinnamic
acid-4-hydroxylase (C4H), r-coumaric acid-3-hydroxylase (C3H), and caffeic acid-o-methytranferase (CAOMT) are involved in the process. Capsaicinoids are formed from vanillylamine and isocapryl-CoA via capsaicinoid synthetases (CS) (Fujiwake et al., 1982; Sukrasno and Yewman, 1993; Curry et al., 1999).
During fruit ripening, capsaicin concentration reaches a maximum. Capsaicin then turns over and degrades to other secondary products (Bernal and Ros Barceló, 1996). Most peroxidase activity occurs in the placenta and the outer layer of pericarp epidermal cells. Histochemical localization was used to determine the location of peroxidase (Bernal et al., 1993a). As determined by gel permeation chromatography, the major oxidative products were 5,5'-dicapsaicin and 4'-O-5-dicapsaicinether (Bernal et al., 1995). Peroxidase activity increased at the time when the concentration of capsaicinoids started to decrease (Contreras-Padilla and Yahia, 1998). It is assumed that peroxidases catalyze capsaicinoid oxidation and play a central role in their metabolism.
The fruits of "Padron" pepper plants given more or less water than control contained higher capsaicin content than well-watered control fruits. Water deficit affects phenylpropanoid metabolism and the pungency of pepper fruits (Quagliotti, 1971; Estrada et al., 1999). PAL, C4H, and CS are involved in capsaicinoid biosynthesis and peroxidase isoenzyme B6 directly affects capsaicin degradation (Bernal et al., 1994a).
The goals of this research were to understand the effect water supply on the amount of capsaicin in hot pep
*Corresponding author. Tel: (04) 2285-1631; E-mail: yusung@dragon.nchu.edu.tw