Bot. Bull. Acad. Sin. (2005) 46: 43-52
LEE et al. Sucrose phosphate synthase from rice
Biochemical characterization of rice sucrose phosphate synthase under illumination and osmotic stress
Mu-Ho Lee, Chien-Chi Yang, Jong-Ching Su, and Ping-Du Lee*
Institute of Microbiology and Biochemistry; Department of Biochemical Science and Technology, College of Life Science, National Taiwan University, Taipei 106, TAIWAN
(Received April 6, 2004; Accepted October 5, 2004)
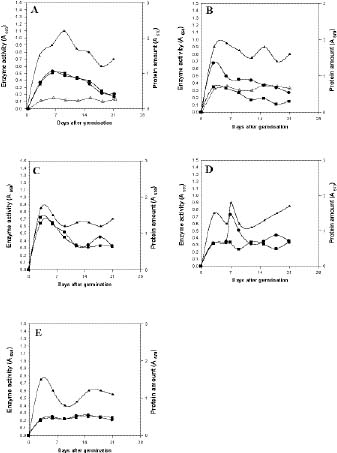
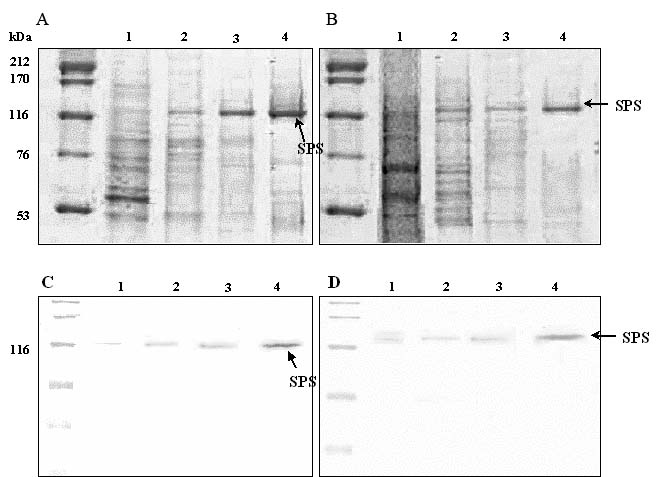
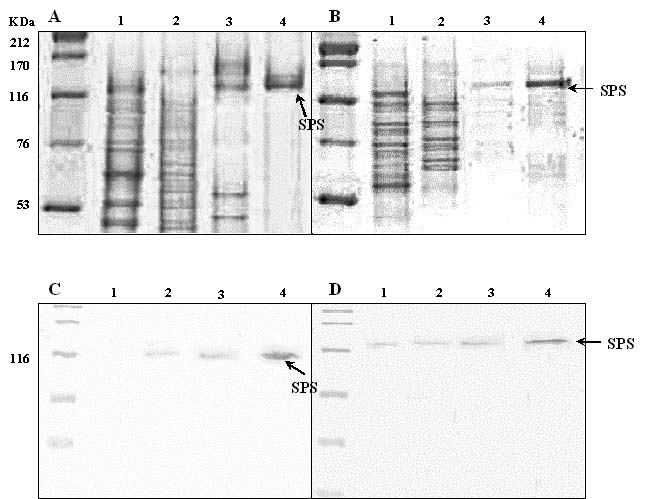
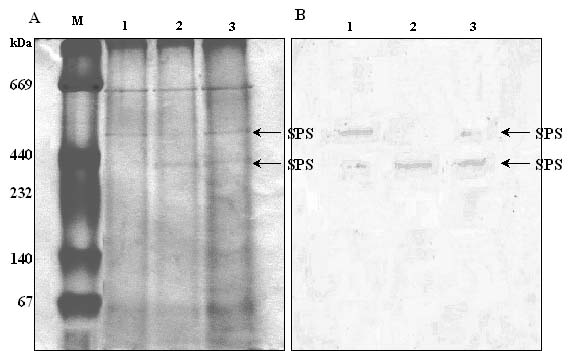
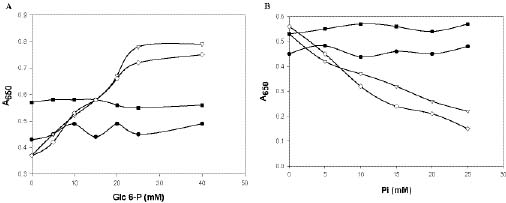
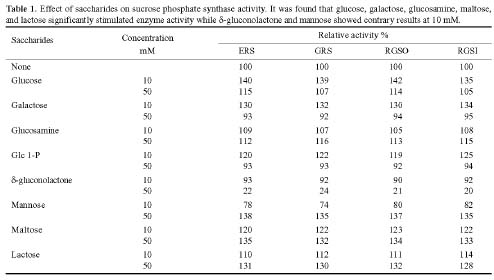
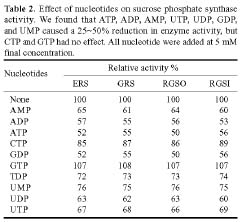
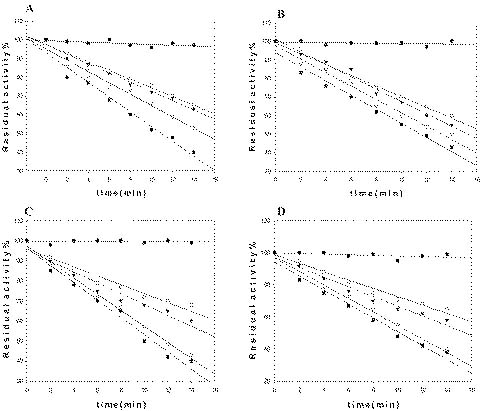
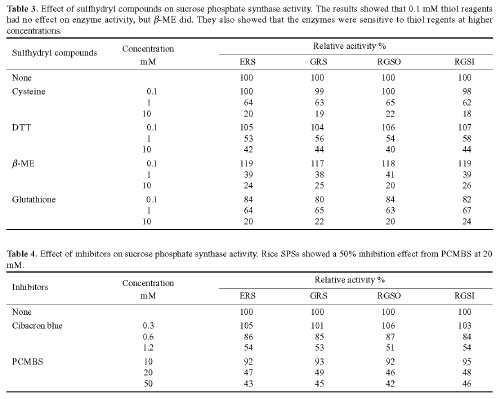
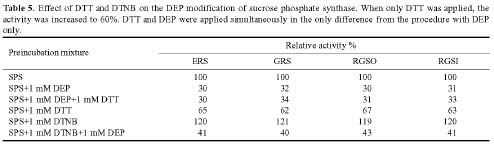
Abstract. Sucrose phosphate synthase (SPS) is one of a number of sucrose-metabolizing enzymes that regulates the sucrose synthesis pathway. SPSs were purified from etiolated rice seedlings (ERS), green rice seedlings (GRS), rice grain suspension cells under osmotic stress (RGSO), and rice grain suspension cells under illumination (RGSI). A native molecular mass of ca. 420 and 520 kDa was found using native-PAGE. The SDS-PAGE analyses revealed SPSs to be homotetramers composed of subunits with a mass of 116-120 kDa. The maximum activity for SPSs was observed on the third day. As far as their biochemical characterization was concerned, the optimum pH of the enzyme reactions lay generally between 6-8, the optimum temperatures between 35-40°C. The ERS and RGSO SPS Km values for Fru 6-P and UDPG were 1.8 and 35 mM, respectively. However, the GRS and RGSI SPS had similar Km values for Fru 6-P and UDPG of 1.5 and 28 mM, respectively. GRS and RGSI SPS activities were allosterically regulated by Glc 6-P (activator) or Pi (inhibitor), but ERS and RGSO SPS had no effect. From their regulations and Km values two enzyme forms (SPS-I and SPS-II) could be discriminated in the rice. SPS-II was induced by illumination, but SPS-I by osmotic stress. All SPSs were activated by Mg2+. The nucleotides AMP, ADP, ATP, UMP, UDP, GDP and UTP inhibited enzyme activity by about 25-50%. Thiol reagents became sensitized to the enzyme activity, but could be restored with DTT or b-ME. Glucose, galactose, glucosamine, maltose, and lactose activated the enzymes and were inhibited by d-gluconolactone and mannose. SPSs were also inhibited by PCMBS, cibacron blue F3G-A, and DEP.
Keywords: Etiolated rice seedlings; Green rice seedlings; Illumination; Isoforms; Osmotic stress; Rice grain suspension cells; Sucrose phosphate synthase.
Abbreviations: DEP, diethylpyrocarbonate; DTNB, 5,5'-dithiobis-2-nitrobenzoic acid; DTT, 1,4-dithiothreitol; ERS, etiolated rice seedlings; GRS, green rice seedlings; RGSI, rice grain suspension cells under illumination; RGSO, rice grain suspension cells under osmotic stress; b-ME, b-mercaptoethanol; PCMBS, p-chloromercuribenzenesulphonic acid; Glc 6-P, glucose 6-phosphate; Fru 6-P, fructose 6-phosphate; Pi, inorganic phosphate; UDPG, uridine 5'-diphosphoglucose.
Introduction
Sucrose is the major stable product of photosynthesis for many plants, and it is the form in which most carbon is transported in phloem vessels from photosynthetic organs into non-photosynthetic organs such as roots, grains, fruits, and tubers (Chris et al., 1999; Rees, 1984). Sucrose accumulates in most plant tissues. This is essential if the plant is to be able to respond effectively to environmental stresses like low temperatures (Guy, 1990), water (Morgan, 1984; Quick et al., 1989; Ramos et al., 1999), and salinity (Balibrea et al., 1997). However, sucrose can also act as a regulator, carbohydrate store, and substrate for biosynthesis and is temporarily preserved in the leaves as a buffer for plants (Chris et al., 1999).
Sucrose synthesis can be catalyzed by the coordination of two enzymes in higher plants: sucrose phosphate
synthase (E.C. 2.4.1.14; SPS) and sucrose phosphate phosphatase (E.C. 3.1.3.24; SPP) (Huber and Huber, 1996). The catalytic reaction is as follows:
SPS is known to be a key regulative enzyme responsible for sucrose synthesis in plants (Stitt et al., 1988; Huber and Huber, 1992; Huber and Huber, 1996). All sucrose-synthesizing organisms contain SPS, which is regulated by several interacting mechanisms, including: (i) Regulation of gene expression (Huber and Huber, 1996), (ii) Covalent modification via reversible phosphorylation (Huber and Huber, 1996), and (iii) Allosteric regulation via metabolites (Doehlert and Huber, 1985).
When purifying SPS in the leaves of plants, it was found that SPS activity changes in coordination with the light cycle (Galtier et al., 1995). In addition, SPS can be activated in spinach leaves under illumination and mannose treatments, but is inhibited under darkness and when inorganic phosphate (Douglas and Huber, 1983) comes in
*Corresponding author. Tel & Fax number: 02-23633610; E-mail: pingdu@ntu.edu.tw