Bot. Bull. Acad. Sin. (2005) 46: 93-98
XU et al. Analysis of citrus intergeneric somatic hybrids
FCM, SSR and CAPS analysis of intergeneric somatic hybrid plants between Changshou kumquat and Dancy tangerine
Xiaoyong XU, Jihong LIU*, and Xiuxin DENG
National Key Laboratory of Crop Genetic Improvement, National Center of Crop Molecular Breeding, Huazhong Agricultural University, Wuhan 430070, P.R. China
(Received July 1, 2004; Accepted December 6, 2004)
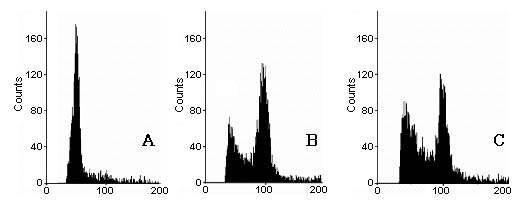
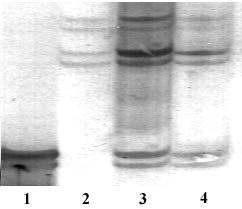
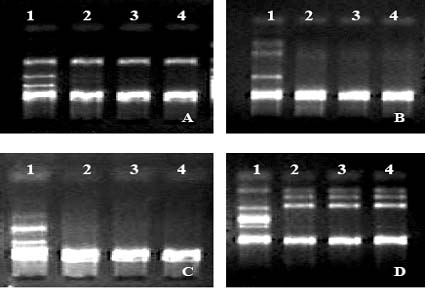
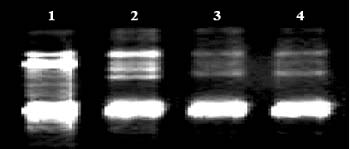
Abstract. Flow cytometry (FCM), simple sequence repeats (SSR) and cleaved amplified polymorphic sequence (CAPS) were used to characterize the ploidy level and the nuclear and cytoplasmic compositions of two intergeneric somatic hybrids of Changshou kumquat (Fortunella obovata), leaf parent, and Dancy tangerine (Citrus reticulata), suspension parent. FCM showed that both somatic hybrids were tetraploids. One out of two SSR primer pairs was able to detect polymorphisms between the fusion parents, and the somatic hybrids showed additive banding profiles, showing them to be allotetraploid hybrids with inherited nuclear DNA from both parents. CAPS showed that 4 and 1 polymorphic loci were obtained for parental chloroplast and mitochondrial genomes, respectively. The somatic hybrid plants had the same banding patterns as the suspension parent for all of the polymorphic chloroplast markers, and their mitochondrial banding patterns approximated those of the suspension parent with loss of some mtDNA fragments. The present research plays a pivotal role in understanding the relationship between nuclear and cytoplasmic compositions and the likely field performance of somatic hybrids in the future.
Keywords: Allotetraploid; Citrus; cpDNA; mtDNA; Somatic hybrids.
Introduction
Somatic hybridization via protoplast fusion paves the way for circumventing some reproductive barriers in citrus traditional breeding, such as polyembryony (nucellar embryos), female and/or male sterility, high degrees of heterozygosity, inconsistencies in flowering period, and sexual incompatibility. Since the first somatic hybrid of trifoliate orange (Poncirus trifoliata) and sweet orange (Citrus sinensis) was produced by Ohgawara et al. (1985), more than 250 combinations of citrus somatic hybrids have been produced worldwide, including sexually incompatible and compatible ones (Grosser et al., 2000; Grosser and Chandler, 2003; Liu et al., 2004). The somatic hybrids hold great potential for citrus cultivar improvement because some of them can be tried as rootstocks, and others can be experimented on for scion cultivars. Several of them have taken on some desirable traits and will be possibly integrated into breeding programs. In addition to producing somatic hybrids with potential for cultivar improvement, somatic hybridization is useful in the study of nuclear-cytoplasmic interactions (nuclear-nuclear, nuclear-cytoplasmic and cytoplasmic-cytoplasmic), owing to its unique advantages over conventional methods, such as the partial transfer of desirable genome (genes) from donor species, the transfer of cytoplasmic factors without changing the nuclear background, and a combination of cytoplasms from different sources, which can not be completed by cross hybridization due to maternal inheritance
of cytoplasmic genomes. Nuclear-cytoplasmic interactions and the final existence fate of the nuclear and cytoplasmic genomes derived from the interactions have certain effects on the growth, development, and field performance of the somatic hybrids. Cheng et al. (2003) reported that abnormal shoot growth of intergeneric somatic hybrids between sweet orange and kumquat was concurrent with loss of mtDNA from one parent. A correlation between mtDNA compositions and field parameters was detected in potato somatic hybrids (Lossl et al., 2000). Studies on nuclear-cytoplasmic interactions depend on reference to detailed somatic hybrid compositions gleaned from morphological, cytological, and genetic markers. Some citrus somatic hybrids have been analyzed using these markers. Previously isozyme, RAPD (randomly amplified polymorphic DNA), and RFLP (restriction fragment length polymorphism) were the main methods used to identify nuclear and cytoplasmic compositions. With the development of new markers, such as SSR (simple sequence repeats) and CAPS (cleaved amplified polymorphic sequence), hybridity identification became rapid and easy. SSR and CAPS have been employed in the verification of some citrus somatic hybrids and in the clarification of the maternal inheritance of some true citrus fruit trees (Liu et al., 2002; Medina-Urrutia et al., 2004; Abkenar et al., 2004; Xu et al., 2004; Guo et al., 2004). In the present paper these two markers, coupled with flow cytometry (FCM), were used to investigate the ploidy level and the nuclear and cytoplasmic constitutions of two intergeneric somatic hybrids of Citrus reticulata and Fortunella obovata with the intention of providing information for further evaluation of their agronomic performance.
*Corresponding author. E-mail: liujihong@mail.hzau.edu.cn