Bot. Bull. Acad. Sin. (2005) 46: 205-215
CHEN and CHEN Ultrastructure of laminar hydathodes
Study on laminar hydathodes of Ficus formosana (Moraceae) I. Morphology and ultrastructure
Chyi-Chuann CHEN and Yung-Reui CHEN*
Institute of Molecular and Cellular Biology, National Taiwan University, Taipei, Taiwan
(Received November 15, 2004; Accepted February 22, 2005)
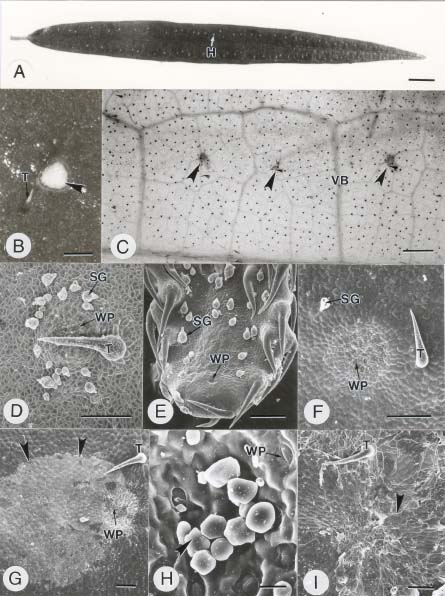
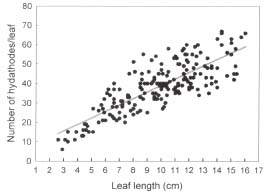
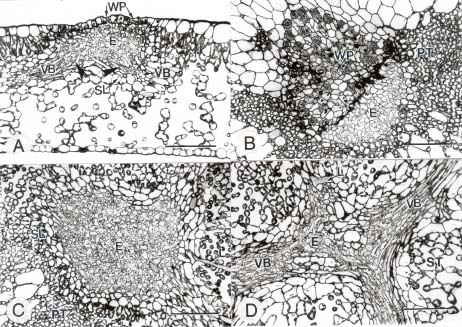
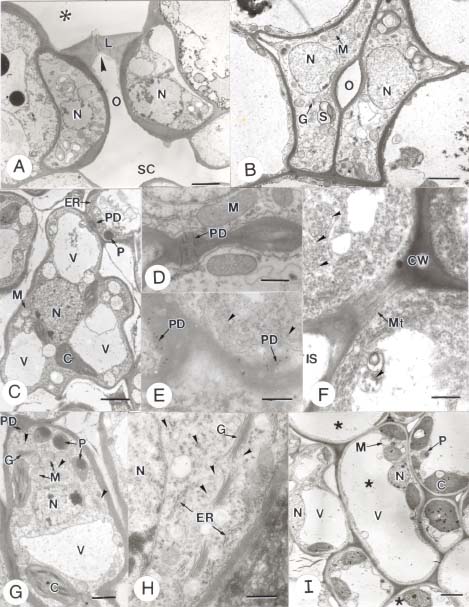
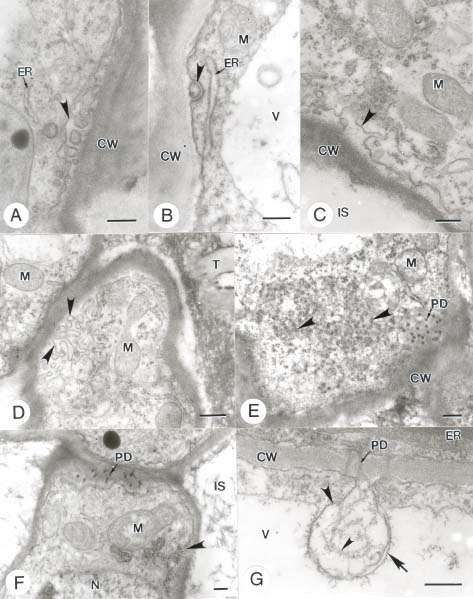
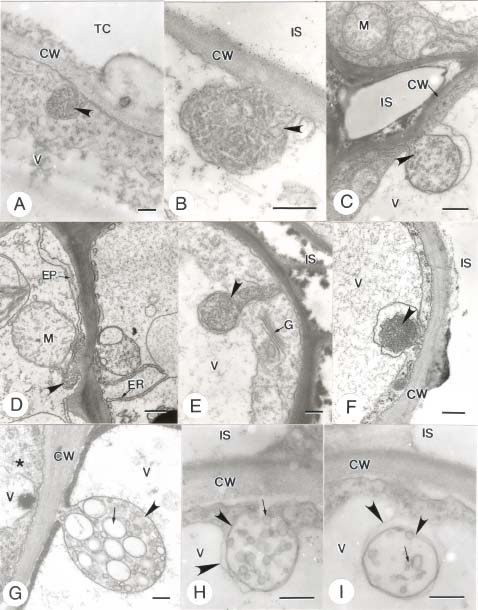
Abstract. In this study, the clearing method, LM, SEM, and TEM were used to examine the morphology and ultrastructures of laminar hydathodes of Ficus formosana Maxim. f. Shimadai Hayata. Morphologically, the laminar-hydathode of F. formosana is complex epithemal type, consisting of water pores, tracheid-ends, epithem cells, and a bounding sheath layer. Water pores are made up of two guard cells, and their pores open permanently. Ultrastructural and cytological data demonstrated that the epithem cell has a dense cytoplasm, numerous mitochondria, an extended ER system, and many small vesicles derived from Golgi bodies. It has proliferate peroxisomes, and their numbers increase with the maturation and aging of the epithem. In addition, abundant plasmodesmata were observed on the contacted cell wall between epithem cells. Variable structures of plasmalemmasome were also observed on the plasma membrane of the epithem. Plasmalemmasome is the result of endocytosis caused by the plasmolysis-deplasmolysis cycle, which is induced by repeating transpiration and guttation day and night. We suggest that the hydathode is an ideal system for studying endocytosis in plants.
Keywords: Endocytosis; Epithem; Ficus formosana Maxim. f. Shimadai Hayata; Laminar hydathodes;Plasmalemmasome; Ultrastructure; Water pores.
Introduction
Guttation is the process by which water exudes from a leaf surface. It differs from transpiration, in which water passes through the stomata in gas form. Guttation is observed in a wide range of vascular plants and ferns. The hydathode is the apparatus of guttation that consists of stomata-like pores in the epidermis and the epithem, which has a large chamber with masses of thin-wall parenchyma cells and a sheath layer surrounding its tissues (Kramer, 1945). In the early studies of hydathodes, Haberlandt (1914) and Lepschkin (1923) divided them into epidermal and epithemal hydathodes. In the former, water secretion is dependent on the metabolic activity of the specialized epidermal cells on the adaxial surface of leaves near the end of the veins, and this is regarded as an active process. For epithemal hydathodes, on the other hand, the process is passive. They respond directly with the water-conducting system of the plant. The water passing through water pores can be observed when the "root pressure" reaches a certain threshold, as a result of an inhibited or a reduced transpiration.
Hydathodes have been found in different dicot families, including Asteraceae (Lersten and Curtis, 1985), Begoniaceae (Brouillet et al., 1987), Rosaceae (Lersten and Curtis, 1982), Balsaminaceae (Elias and Gelband, 1977), Salicaceae (Curtis and Lersten, 1974), Urticaceae (Smith and
Watt, 1986; Lersten and Curtis, 1991), Crassulaceae (Rost, 1969), and Moraceae (Lersten and Peterson, 1974), as well as in cereal plants (Maeda and Maeda, 1987, 1988; Dieffenbach et al., 1980) and pteridophytes (Sperry, 1983). Most of the hydathodes can be found at the tip, on the margin, and/or over the entire surface of leaf. In a small number of species, hydathodes are found on leaf hypodermis (Fahn, 1979). In a few cases, they can also be found in other organs. For example, hydathode is found on the grape tendril (Vitis vinifera) (Tucker and Hoefert, 1968). They are also found in certain submerged aquatic plants (Pedersen et al., 1997).
The current study focuses on a particular type of hydathode, the so-called "laminar hydathodes." To our knowledge, three dicot familiesi.e., Moraceae, Urticaceae, and Crassulaceaehave this type hydathode distribution (Lersten and Curtis, 1991). On the hydathodes of Ficus, in Moraceae, little data have ever been published. For example, Molisch (1916) described the anatomy of the hydathode in F. javanica, and Lersten and Peterson (1974) later illustrated the structure of hydathodes in F. diversifolia using light microscopy. A detailed investigation into the ultrastructure of the laminar hydathodes in Ficus has not yet been completed. The aims of this study were to present the ultrastructural data of the adaxial-laminar hydathodes in F. formosana Maxim. f. Shimadai Hayata through the clearing method, light microscopy, scanning electron microscopy, and transmission electron microscopy. It is intended to serve as a platform for further studies on the function of laminar hydathodes.
*Corresponding author. Fax: 02-3366-2478; E-mail: yrc@ntu.edu.tw