|
tance genes in transgenic tobacco plants. Crop Prot.
9: 351-354.
Bown, D.P., H.S. Wilkinson, and J.A. Gatehouse. 1997.
Differentially regulated inhibitor sensitive and insensitive protease genes
from the phytophagous insect pest, Helicoverpa armigera, are members of
complex multigene families. Insect Biochem. Mol. Biol. 27: 625-638.
Broadway, R.M. 1996. Dietary proteinase inhibitors alter
complement of midgut proteases. Arch. Insect Biochem. Physiol. 32:
39-53.
Bryant, J., T.R. Green, T. Gurusaddaiah, and C.A. Ryan.
1976. Proteinase inhibitor II from potatoes: isolation and characterization of
its protomer components. Biochemistry 15: 3418-3424.
Brzin, J. and M. Kidric. 1995. Proteinases and their
inhibitors in plants: role in normal growth and in response to various stress
conditions. Biotechnol. Genet. Eng. Rev. 13: 420-467.
Burgess, E.P.J. and A.M.R. Gatehouse. 1997. Engineering for
insect pest resistance. In B.D. McKersie and D.C.W. Brown (eds.),
Biotechnology and the Improvement of Forage Legumes. New York: CAB
International, pp. 229-258.
Cai, D., T. Thurau, Y. Tian, T. Lange, K.W. Yeh, and C.
Jung. 2003. Sporamin-mediated resistance to beet cyst nematodes (Heterodera
schachtii Schm.) is dependent on trypsin inhibitory activity in sugar beet
(Beta vulgaris L.) hairy roots. Plant Mol. Biol. 51:
839-849.
Callihan, R.H., J.C. Ojala, L.C. Haderlie, and D.W. Kidder.
1990. Nightshade: biology and control in cropland of the Pacific Northwest.
Pac. Northwest. Ext. Pub. 352: 1-6.
Carbonero, P., J. Royo, I. Diaz, F. Garcia-Maroto, E.
Gonzalez-Hidalgo, C. Gutierez, and P. Cassanera. 1993. Workshop on Engineering
Plants Against Pests and Pathogens, In G.J. Bruening, F. Garcia-Olmedo,
and F.J. Ponz (eds.), January, Instituto Juan March de Estudios
Investigaciones, Madrid, Spain, pp. 1-13.
Charity, J.A., M.A. Anderson, D.J. Bittisnich, M.
Whitecross, and T.J.V. Higgins. 1999. Transgenic tobacco and peas expressing a
proteinase inhibitor from Nicotiana alata have increased insect
resistance. Molecular Breed. 5: 357-365.
Chi, G.-L., D.G.. Barfield, G..-E. Sim, and E.-C. Pua.
1990. Effect of AgNO3 and aminoethoxyvinyl- glycine on in
vitro shoot and root organogenesis from seedling explants of recalcitrant
Brassica genotypes. Plant Cell Rep. 9: 195-198.
Chiche, L, A. Heitz, J.C. Gelly, J. Gracy, P.T. Chau, P.T.
Ha, J.F. Hernandez, and D. Le-Nguyen. 2004. Squash inhibitors: from structural
motifs to macrocyclic knottins. Curr. Protein Pept. Sci. 5:
341-349.
Choi, D., J.-A. Park Y.S. Seo, Y.J. Chun, and W.T. Kim.
2000. Structure and stress-related expression of two cDNAs encoding proteinase
inhibitor II of Nicotiana glutinosa L. Biochim. Biophys. Acta.
1492: 211-215.
Chrispeels, M.J. and B. Baumgartner. 1978. Trypsin
inhibitor in mung bean cotyledons: purification, characteristics, subcellular
localization, and metabolism. Plant Physiol. 61: 617-623.
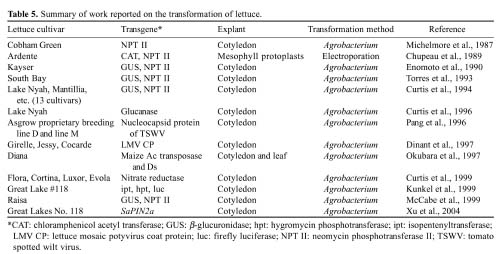
Chupeau, M.C., C. Bellini, P. Guerche, B. Maisonneuve, G.
Vastra, and Y. Chupeau. 1989. Transgenic plants of lettuce (Lactuca
sativa) obtained through electroporation of protoplasts. Bio/Technol.
7: 503-508.
Cipriani, G., D. Michaud, F. Brunelle, A. Golmirzaie, and
D.P.
|
|
Zhang. 1999. CIP Program Report 1997-98, 1999, 271-277.
Clark, A.M., K.R. Jacobsen, D.E. Bostwick, J.M.
Dannenhoffer, M.I. Skaggs, and G.A. Thompson. 1997. Molecular characterization
of a phloem-specific gene encoding the filament protein, phloem protein 1
(PP1), from Cucurbita maxima. The Plant J. 12: 49-61.
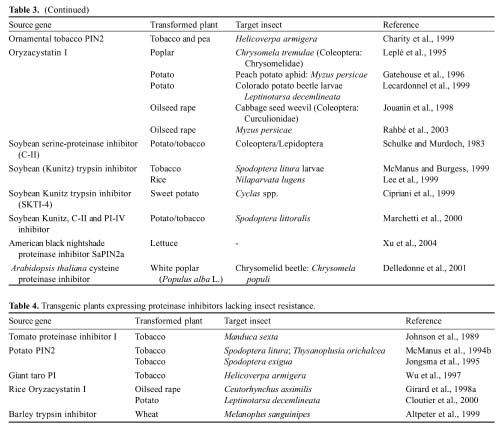
Cloutier, C., C. Jean, M. Fournier, S. Yelle, and D.
Michaud. 2000. Adult Colorado potato beetles, Leptinotarsa decemlineata
compensate for nutritional stress on oryzacystatin I-transgenic potato plants
by hypertrophic behavior and over-production of insensitive proteases. Arch.
Insect Biochem. Physiol. 44: 69-81.
Connors, B.J., N.P. Laun, C.A. Maynard, and W.A. Powell.
2002. Molecular characterization of a gene encoding a cystatin expressed in
the stems of American chestnut (Castanea dentata). Planta 215:
510-514.
Creemers, J. 2002. Proteolytic enzymes as therapeutic
targets - keystone symposium. I Drugs 5: 216-219.
Curtis, I.S., C. He, R. Scott, J.B. Power, and M.R. Davey.
1996. Genomic male sterility in lettuce, a baseline for the production of
F1 hybrids. Plant Sci. 113: 113-119.
Curtis, I.S., J.B. Power, A.M.M. de Laat, M. Caboche, and
M.R. Davey. 1999. Expression of a chimeric nitrate reductase gene in
transgenic lettuce reduces nitrate in leaves. Plant Cell Rep. 18:
889-896.
Curtis, I.S., J.B. Power, N.W. Blackhall, A.M.M. de Laat,
and M.R. Davey. 1994. Genotype- independent transformation of lettuce using
Agrobacterium tumefaciens. J. Exp. Bot. 45:
1441-1449.
De Leo, F., M. Volpicella, F. Licciulli, S. Liuni, R.
Gallerani, and L.R. Ceci. 2002. PLANT-PIs: a database for plant protease
inhibitors and their genes. Nucl. Acids Res. 30: 347-348.
De Leo, F. and R. Gallerani. 2002. The mustard trypsin
inhibitor 2 affects the fertility of Spodoptera littoralis larvae fed
on transgenic plants. Insect Biochem. Mol. Biol. 32: 489-496.
De Leo, F., M. Bonadè-Bottino, L.R. Ceci, R. Gallerani, and
L. Jouanin. 2001. Effects of a mustard trypsin inhibitor expressed in
different plants on three lepidopteran pests. Insect Biochem. Mol. Biol.
31: 593-602.
De Leo, F., M.A. Bonade-Bottino, L.R. Ceci, R. Gallerani,
and L. Jouanin. 1998. Opposite effects on spodoptera littoralis larvae of high
expression level of a trypsin proteinase inhibitor in transgenic plants. Plant
Physiol. 118: 997-1004.
Delledonne M., G. Allegro, B. Belenghi, A. Balestrazzi, F.
Picco, A. Levine, S. Zelasco, P. Calligari, and M. Confalonieri. 2001.
Transformation of white poplar (Populus alba L.) with a novel
Arabidopsis thaliana cysteine proteinase inhibitor and analysis of insect pest
resistance. Mol. Breed. 7: 35-42.
Dinant, S., B. Maisonneuve, J. Albouy, Y. Chupeau, M.C.
Chupeau, Y. Bellec, F. Gaudefroy, C. Kusiak, S. Souche, C. Robaglia, and H.
Lot. 1997. Coat protein gene-mediated protection in Lactuca sativa
against lettuce mosaic potyvirus stains. Molecular Breed. 3: 75-86.
Ding, L.-C., C.-Y. Hu, K.-W. Yeh, and P.-J. Wang. 1998.
Development of insect-resistant transgenic cauliflower plants expressing the
trypsin inhibitor gene isolated from local sweet potato. Plant Cell Rep.
17: 854-860.
Doares, S.H., J. Narvaez-Vasquez, A. Conconi, and C.A.
Ryan. 1995. Salicylic acid inhihits synthesis of proteinase inhibi
|
|