Bot. Bull. Acad. Sin. (2005) 46: 339-345
LIN et al. Genome organization and relationships of Phalaenopsis
Genome organization and relationships of Phalaenopsis orchids inferred from genomic in situ hybridization
Chih-Chung LIN1, Yao-Huang CHEN2,3, Wen-Huei CHEN2,4, Chi-Chang CHEN1, and Yen-Yu KAO1,5,*
1Department of Botany, National Taiwan University, Taipei 106, Taiwan, ROC
2Department of Horticulture, Taiwan Sugar Research Institute, Tainan 701, Taiwan, ROC
(Received April 21, 2005; Accepted July 13, 2005)
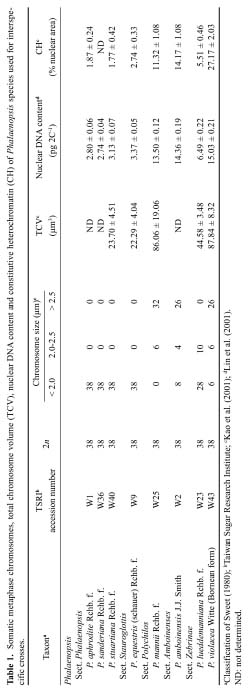
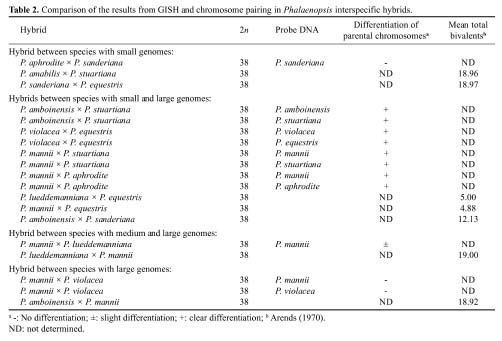
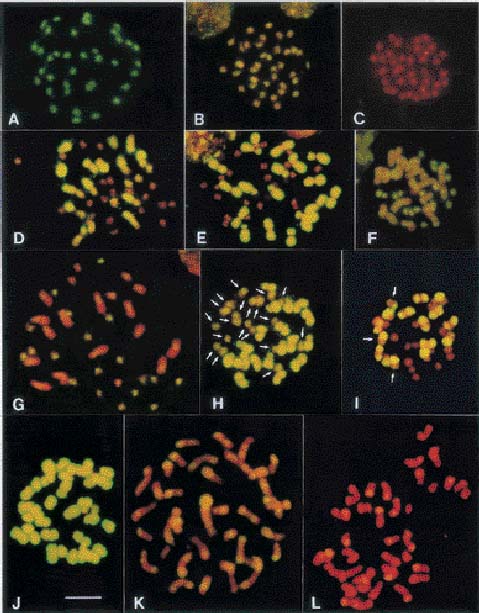
Abstract. All Phalaenopsis species have the same chromosome number (2n = 2x = 38), but the sizes of their genomes vary markedly. In this study, we investigated genome organization and relationships of Phalaenopsis species by genomic in situ hybridization (GISH) of seven interspecific hybrids derived from crosses between species with similar or dissimilar sizes of genomes. In the hybrid P. aphrodite × P. sanderiana, in which both parents possess small genomes, the two parental chromosome sets could not be distinguished by the strength and distribution of hybridization signals. Similar results were obtained from the hybrid P. mannii × P. violacea, in which both parents have large genomes. These results suggest that the two parents of these hybrids have similar genomes. In hybrids in which one parent possesses a large and the other parent a small genome, such as P. amboinensis × P. stuartiana, the two parental chromosome sets could readily be distinguished by GISH with or without the application of blocking DNA. Examination of hybridization signals on chromosomes revealed that species with large genomes have much more repetitive sequences, both in type and amount; however, species with small genomes do have their own specific sequences. In general, genome relationships obtained from GISH are in agreement with those from traditional genome analysis of other investigators. This study demonstrates that GISH is a useful tool for investigating genome organization and relationships of plant species, especially when analysis of meiotic behavior is technically difficult.
Keywords: Genomic in situ hybridization; Genome organization; Genome relationship; Phalaenopsis.
Introduction
Genome analysis provides valuable information about species relationships and, therefore, plays an important role in plant breeding programs. Among the traditional methods of genome analysis, studying the meiotic chromosome pairing in F1 hybrids is most reliable and has been frequently used because it gives direct evidence for genome homology between parental species (Singh, 2003).
The genus Phalaenopsis (Orchidaceae) comprises 45 to 63 species and has a wide geographic distribution, ranging from the Himalayas of northern India through Southeast Asia to northern Australia (Sweet, 1980; Christenson, 2001). Species and hybrids in this genus are of high value in floriculture because of their graceful and long-lasting flowers. Although interspecific and intergeneric hybrid
ization has long been used for breeding superior cultivars in Phalaenopsis orchids, so far only one report has investigated meiotic chromosome pairing in F1 hybrids (Arends, 1970). Analysis of chromosome pairing in Phalaenopsis is difficult because: (1) plants are slow-growing and require about two to three years to reach maturity; (2) each plant produces very few flowers, hindering collection of sufficient microsporocytes at the right stages for analysis; (3) microsporocytes are enclosed in a thick callose wall, which hampers stain penetration; and (4) meiotic chromosomes cannot be spread well due to clumping and stickiness.
Two molecular techniques, genomic Southern hybridization and genomic in situ hybridization (GISH), have been developed (Schwarzacher et al., 1989; Anamthawat-Jónsson et al., 1990) which may complement the method of genome analysis by meiotic chromosome pairing. Both techniques use total genomic DNA as a probe, but the hybridization targets are different. In genomic Southern hybridization the target is DNA on Southern blots while in GISH the target is chromosomes prepared from mitotic or meiotic cells. Both techniques detect hybridization of repetitive sequences (Anamthawat-Jónsson et al., 1990), which constitute the main source of genome-sized variation between related species (Flavell, 1982; Kubis et al., 1998).
3Present address: Crop Improvement Division, Tainan District Agricultural Research & Extension Station, Tainan 712, Taiwan, ROC
4Present address: Department of Life Science, National University of Kaohsiung, Kaohsiung 811, Taiwan, ROC
5Present address: Institute of Molecular and Cellular Biology, National Taiwan University, Taipei 106, Taiwan, ROC
*Corresponding author. E-mail: yykao@ntu.edu.tw; +886-2-33662486; Fax: +886-2-33662478.