Bot. Bull. Acad. Sin. (2005) 46: 347-353
ZHANG et al. Abscisic acid affects self-thinning
Sensitivity of response to abscisic acid affects the power of self-thinning in Arabidopsis thaliana
Hao ZHANG, Gen-Xuan WANG*, Zhi-Qiang LIU, Zhu-Xia SHEN, and Xing-Zheng ZHAO
State Key Laboratory of Plant Physiology & Biochemical, College of Life Sciences, Zhejiang University, Hangzhou, Zhejiang Province, 310029, P. R. China
(Received November 12, 2004; Accepted May 11, 2005)
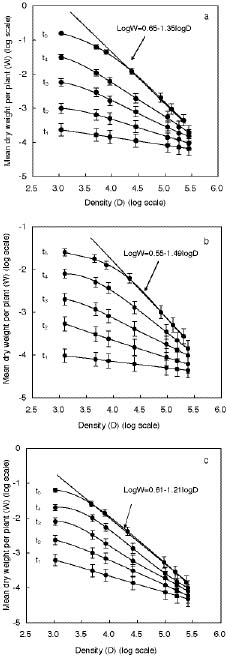
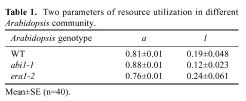
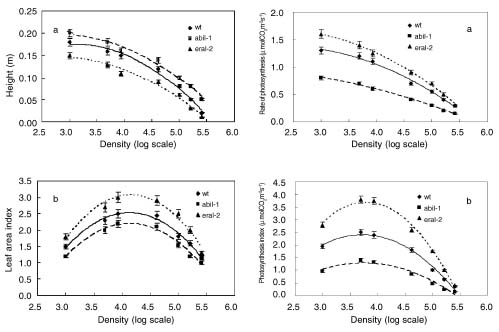
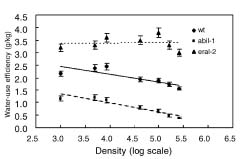
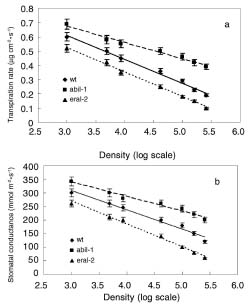
Abstract. The effect of sensitivity of abscisic acid (ABA) on the power of self-thinning was studied with two Arabidopsis mutants (abi1-1, insensitive to ABA and era1-2 hypersensitive to ABA). The experimental results showed that the self-thinning power of abi1-1 (-1.49) was smaller than that of wild type (-1.35), and the self-thinning power of era1-2 (-1.21) was greater than that of wild type. Two parameters of resource utilization (l mean height from root to leaf; a total area of leaves) were more advantageous to era1-2 than to abi1-1 under density stress. Physiological indices of light use (photosynthetic rate, photosynthesis index PSI), water loss (transpiration rate, stomata area index SAI), and water-use efficiency (photosynthetic rate/ transpiration rate) of abi1-1 and era1-2 were consistent with the resource utilization parameters of these Arabidopsis mutants. It can be concluded that the different self-thinning power of the two Arabidopsis mutants resulted from their different resource utilization (such as light and water use) in response to density stress.
Keyword: Abscisic acid; Arabidopsis thaliana; Photosynthesis; Resource utilization; Self-thinning.
Abbreviation: ABA, abscisic acid; LAI, leaf area index; PSI, photosynthesis index; SAI, stomatal area index.
Introduction
The self-thinning law describes variation in population density with body size in the ecological community (Yoda et al., 1963). The -3/2 self-thinning law is described by Formula 1:
logW = logK + blogD (1)
where W and D were the average weight and real density of surviving individuals, respectively (Yoda et al., 1963). Also, K and b were the constant and the power of self-thinning, respectively. Many ecological scientists have devoted themselves to research on the self-thinning law for the past 30 years, and none of this research has disproven the -3/2 or -4/3 self-thinning law (White and Harper, 1970; Harper, 1977; White, 1980, 1981, 1985; Westoby, 1984; Hutching, 1983; Dewar, 1993). Moreover, the power of self-thinning was extended from -1 to -2, according to the relationship between the weight and metabolic rate of animal and microbe (West et al., 1997, 1999b, 2001). Results also indicated that the power of self-thinning phenomena is usually regulated by abiotic or biological factors (Yoda et al., 1963; Weller, 1987a). Some abiotic factors, such as light, water, nutrition and temperature, can affect the power of self-thinning in plant communities directly (Thomas and Weiner, 1989; Morris, 1999; Callaway,
2002). Additionally, gene expression, intracellular signaling, and hormone response may also affect the power of self-thinning in plant communities indirectly.
The power of the self-thinning law should be mediated by the rate of resource utilization, which is in proportion to the physiological metabolism of the plant (Equist et al., 1998; Brian et al., 2003; Jorg et al., 2003). The model of the four dimensions of life showed that all unicellular and multicellular organisms have either virtual or real fractal-like distribution networks for the internal transport of metabolites, thereby endowing them with a "fourth spatial dimension" (Equist et al., 1998; West et al., 1999b). These networks are purported to maximize metabolic capacity and efficiency by maximizing the available surface area for absorption of limiting resources from the environment, yet minimizing transport distance and time. In the N (N ³ 3) dimension, the power of the self-thinning law should be modulated by parameters of resource utilization, such as l and a, as in Equation 2:
![]()
where ![]() represents the power of self-thinning; 1 is the
internal distance associated with the network of fractal plant and is
negatively correlated with resource utilization rate; and a is
effective surface area or fractal dimension and positive to resource
utilization . The sum of l and a is 1; the definition of W and D
are the same as in Formula 1.
represents the power of self-thinning; 1 is the
internal distance associated with the network of fractal plant and is
negatively correlated with resource utilization rate; and a is
effective surface area or fractal dimension and positive to resource
utilization . The sum of l and a is 1; the definition of W and D
are the same as in Formula 1.
*Corresponding author. E-mail: wanggx@zju.edu.cn; Tel and Fax: +86-571-86971083.