Bot. Bull. Acad. Sin. (2005) 46: 355-361
LI The cytogeography of Aster ageratoides var. laticorymbus
The cytogeography of Aster ageratoides var. laticorymbus (Asteraceae), a polyploid complex endemic to China
Wei-Ping LI*
College of Life Sciences, Hunan Normal University, Changsha 410081, P.R. China
(Received July 2, 2004; Accepted June 2, 2005)
Abstract. Aster ageratoides var. laticorymbus is distrubuted from southwestern China to eastern China. Population sampling across its whole distribution area was made in its twenty-six populations. Chromosome numbers of all the populations were investigated, and six populations, representative of various ploidy levels, were analyzed karyotypically for the first time. The results show that this variety is a polyploid complex comprised of 2x, 4x and 6x. The populations are mostly hexaploid (2n = 6x = 54) and occupy an extensive area from southwestern China to eastern China while the diploid (2n = 2x = 18) and the tetraploid (2n = 4x = 36) are less frequent and limited to narrower regions of the Yunnan-Guizhou Plateau and the transitional belts from the plateau to adjacent areas. Based on the cytogeographical distribution of the complex, the analysis of its karyotypes, its morphological variations related to geography and a previous report, some hypothesis are made: (1) the polyploids of the complex might be autopolyploids, and the tetraploid might have originated independently twice; (2) in terms of some karyotypical parameters, different ploidy levels of the complex evolved at different speeds; (3) the Yunnan-Guizhou Plateau is the diversity center and the origin center of the complex, and the place from which the hexaploid dispersed eastwards to eastern China; (4) the distribution pattern that the hexaploid is dominating might have been formed by the competitive exclusion and its high capability to disperse and occupy new habits; and (5) the variety is a relative young complex.
Keywords: Aster ageratoides var. laticorymbus; Dispersal route; Distribution pattern; Karyotype; Polyploid complex; Yunnan-Guizhou Plateau.
Introduction
Aster ageratoides Turcz., a perennial herb of Asteraceae (Compositae), is widely distributed from northeastern Asia to southeastern Asia, but with its major diversity center in China (Ling and Chen, 1985; Li, 2002). This species has a complex taxonomic history and is reputed to be a difficult taxonomical subject (Huziwara, 1957; Ling and Chen, 1985; Ito and Soejima, 1995; Soejima and Peng, 1998; Soejima et al., 1999; Li, 2002). According to Ling and Chen (1985), it comprises 11 varieties in China. However, the taxonomic status of these taxa is quite controversial, and some are very difficult to distinguish from each other (Ling and Chen, 1985; Soejima et al., 1999; Li, 2002). The taxonomic confusion in this difficult species comes from the fact that very little is known of its diversity and variation in morphology and cytology (Soejima et al., 1999; Li, 2002).
Aster ageratoides var. latiocorymbus (Vant.) Hand.-Mazz. is endemic to China (Handel-Mazzetti, 1938; Ling and Chen, 1985; Li, 2002). It is characterized by poly-branched stem, oblong-lanceolate or ovate-lanceolate middle leaves, narrow phyllaries with green top, and white ray florets, the narrow phyllaries being the most important diagnostic character (Handel-Mazzetti, 1938; Ling and Chen, 1985; Soejima et al., 1999). This variety is of eco
logical importance due to its participation in the revegetation of degraded or newly open habitats (Li, 2002). Chen et al. (1992a, 1992b) reported for the first time that the variety laticorymbus has two cytotypes (4x = 36; 6x = 54) in Hunan Province and Guangxi Province. Unfortunately, mistakes in identifying specimens impaired the excellence of their work (Li, 2002). The only other report of cytology of the variety was from Soejima et al. (1999), who documented three cytotypes (2x = 18; 4x = 36; 8x = 71, 72) of this variety in the Zhaotong area, northeastern Yunnan.
While working on biosystematic studies on Aster ageratoides, the author made extensive field observations and sampling of 73 native populations in 20 provinces of China between 2000 and 2002. As a part of the work, A. ageratoides var. laticorymbus was investigated in cytogeography and morphology. This study reports the current distribution pattern of three cytotypes of var. laticorymbus, and then discusses its evolution of karyotype, its origin, dispersal route, the formation mechanism of current distribution pattern, and the age of the A. ageratoides var. laticorymbus complex.
Materials and Methods
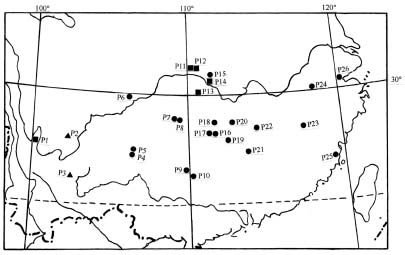
Field observations and sampling were made in 73 populations of Aster ageratoides, of which 26 populations (Table 1; Figure 1) from 11 provinces belonged to A. ageratoides var. laticorymbus according to Li (2002). For each population, more than ten plants were transplanted to
*Corresponding author. E-mail: lwp@hunnu.edu.cn; Tel: 86-731-8871052; Fax: 86-731-8883310.